
Papionina, Burnett, 1828
|
publication ID |
https://doi.org/ 10.5252/geodiversitas2023v45a5 |
|
publication LSID |
lsid:zoobank.org:pub:CB66D102-090F-4AE7-89E8-D83E9024718B |
|
DOI |
https://doi.org/10.5281/zenodo.7822990 |
|
persistent identifier |
https://treatment.plazi.org/id/0277E376-FFFB-D477-FBDA-B3BB672FF58B |
|
treatment provided by |
Felipe |
|
scientific name |
Papionina |
| status |
|
Papionina indet.
MATERIAL EXAMINED. — Isolated teeth. BPB 14 ( Fig. 18 View FIG AC; Tables 6 View TABLE , 7 View TABLE ); BPB 15 ( Fig. 18 View FIG D-F; Tables 6 View TABLE , 7 View TABLE ); BPB 27 ( Fig. 18 View FIG G-H). —– Postcranial bones. BPB 5 ( Fig. 20A, B View FIG ); BPB 6 ( Fig. 20C, D View FIG ).
DESCRIPTION
Isolated teeth
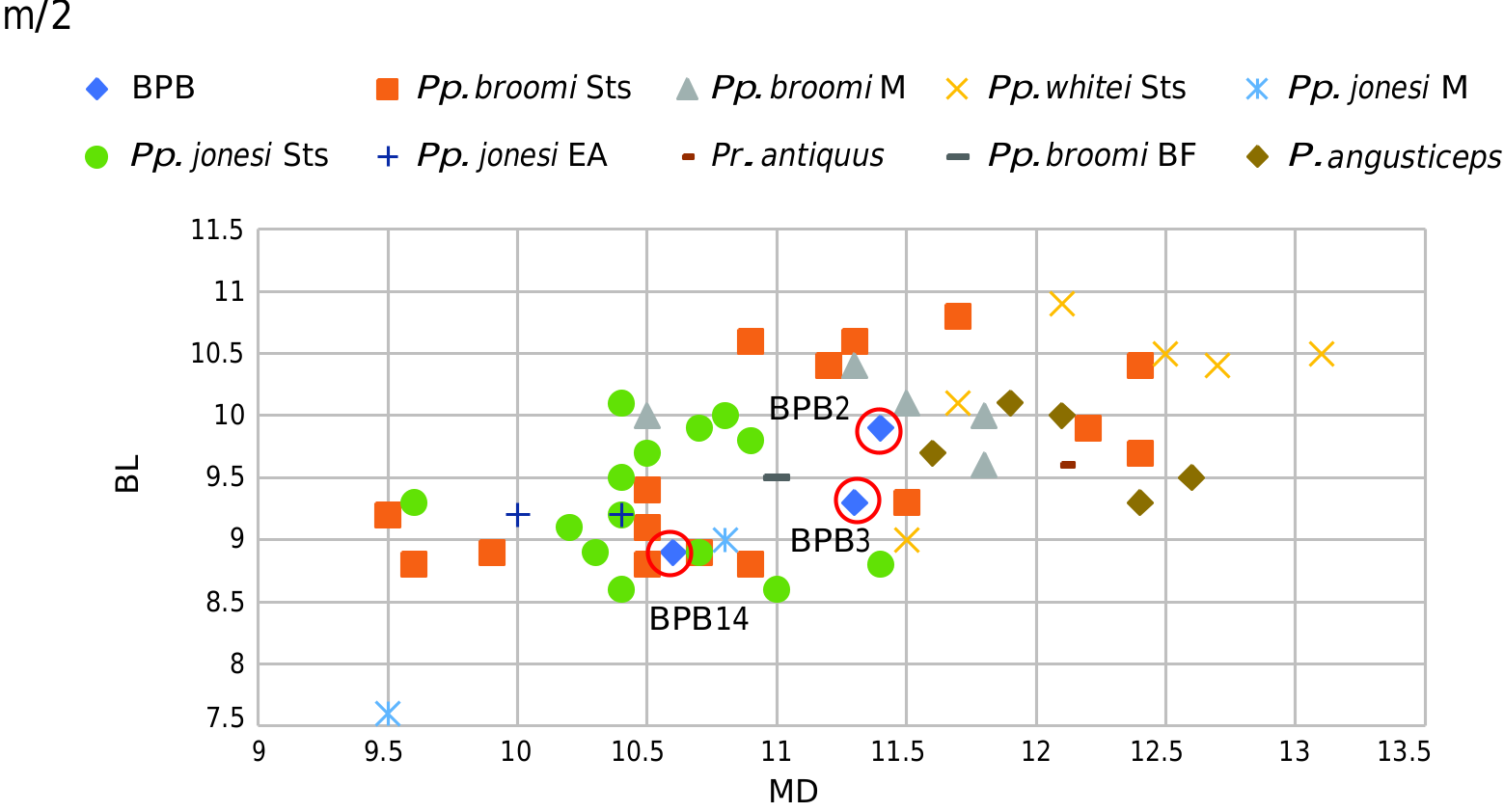
BPB 14 ( Fig. 18 View FIG A-C; Tables 6 View TABLE , 7 View TABLE ). This isolated lower molar is probably a left m/2. The mesial contact surface is bigger and located more buccally than the distal one which is located more centrally. The enamel is thick and the buccal cusps are more worn than the lingual cusps. The protolophid and the hypolophid are low. The protoconid is the largest and the most deeply worn cusp. The metaconid is located slightly more mesially than the protoconid. The median clefts are narrow. The beginning of the mesial and distal roots is preserved. The anatomical features of this tooth correspond to the molars of papionines. BPB 14 is smaller than the m/2 of BPB 2 and BPB 3 but enters into the range variation of the m/2 of Pp. broomi ( Tables 6 View TABLE , 7 View TABLE ; Fig. 12 View FIG ).
BPB 15 ( Fig. 18 View FIG D-F; Tables 6 View TABLE , 7 View TABLE ). The crown of this right p/4 is well preserved and is unworn. The roots are fragmentary. The protoconid and metaconid are close and clearly separated by a V-shaped transverse crest. These two cuspids are moderate salient from the crown and their apex is very slightly worn. The protoconid is slightly lower and is the most prominent cuspid. In buccal view ( Fig. 18F View FIG ), there is a slight mesio-buccal flange but also a deep distal and a mesial buccal cleft. The mesial fovea is small and moderately deep. The talonid is developed and high with a deep distal basin edged by a well-developed distal cingulum. The hypoconid is developed but narrower than the mesial part of the tooth. There is a small entoconid. The prehypocristid and the pre-entoconid are very short. There is a split on the lingual edge of the crown between the prehypocristid and the postprotocristid and between the pre-entoconid and the post-entoconid. The morphology of BPB 15 corresponds to a p/4 of papionines. The size of this premolar enters more into the range of variation of Pp. whitei than that of Pp. jonesi ( Tables 6 View TABLE , 7 View TABLE ; Fig. 19 View FIG ).
BPB 27 ( Fig. 18G, H View FIG ). This small tooth should be a left di/1 and the apex of the root is broken (maximum height: 10.6 mm). The crown is very worn and presents an isosceles triangular outline (MD: 3 mm, BL: 3.9). The enamel is only preserved on the buccal surface (preserved height: 3.5 mm). The root is well preserved except for the apex (preserved height: 7.3 mm). The root is slightly curved. The buccal surface of the root is concave mesio-distally and the lingual one is slightly convex. The morphology of this incisor looks like some juvenile specimens of PlioPleistocene papionines.
Post-cranial bones.
BPB 5 ( Fig. 20A, B View FIG ). This left patella is well preserved except for the disto-inferior part and the median side which have lost some superficial bone (maximum length: 18.2 mm; breadth: 12.9 mm; thickness: 8.3 mm). This patella presents a general morphology similar to WP 3 (a right patella from Waypoint 160, Bolt’s Farm [ Gommery et al. 2008b]), less elongated than in Papio ursinus . In anterior view ( Fig. 20A View FIG ), the medial part is more convex and shorter medio-laterally than the lateral part. In posterior view ( Fig. 20B View FIG ), the medial part of the articular facet is longer proximo-distally and shorter medio-laterally than the lateral part, and as well more concave proximo-distally. The distal apex is low, broad and has a rounded aspect with a groove posteriorly as in WP 3 (corresponding to the insertion of the lateral and medial Vasti and the M. rectus femoris ). Despite the damage posteriorly, the proximal part of the patella is more elongated proximo-distally than the distal part. The morphology of this patella seems to suggest a mixed locomotor behaviour (terrestrial and arboreal).
BPB 6 ( Fig. 20C, D View FIG ). This fossil is a fragment of a right talus (length: 24.4 mm, breadth: 11 mm). Only a part of plantar surface is preserved with the head (Caput tali) and the neck (Collum tali), as well as the distal part of the body of the talus (Corpus tali) ( Fig. 20C View FIG ). The neck is broad relatively to its length (length 14.7 mm; breadth: 10.8 mm). On the head, the plantar part of the articular surface for the navicular is preserved and presents a slightly convex outline. The anterior and medial calcaneal articular surfaces are well preserved. These form a large proximo-distally elongated rectangle and occupy a large part of the surface along the medial edge of the talar neck. This morphology is typical of cercopithecines and differs from those of extant colobines where the two surfaces are distinct ( Olivier & Fontaine 1957; Strasser 1988; Gilbert et al. 2010) but not in Paracolobus mutiwa ( Anderson 2021) . Pina et al. (2011) have observed that the anterior calcaneal surface is more laterally developed in some cercopithecines such as Lophocebus and Miopithecus but not in Papio and many species of Macaca . In BPB 6, this surface is not developed laterally and is more similar to Papio . This morphology of the anterior and the medial articular surfaces is more related to weak rotation of the head of the talus as is usual in cercopithecines and different from the stronger rotation observed in colobines ( Strasser 1988). Laterally, the fovea of the anterior astragalo-calcaneal ligament is moderately deep. The talar sulcus (Sulcus tali) is narrow and shallow laterally but broader and deeper medially below the groove for flexor hallucis longus.
The posterior calcaneal facet is preserved in its distal part. This facet seems deeply concave latero-distally to medio-proximally as in Papio . The junction with the groove for flexor hallucis longus is very salient. This groove has lost some superficial bone and is narrow. There is no swelling distally to the groove for the flexor hallucis longus as in Papio , Cercocebus and Cercopithecus ( Strasser 1988; Gilbert et al. 2010).
BPB 6 is very damaged in dorsal view ( Fig. 20D View FIG ). Medially, a small fragment of the tibial malleolar facet is preserved distally and shows the contact with the medial calcaneal articular surface as in Cercocebus ( Strasser 1988) and in Papio but not in extant colobines. A fragment of the medial tubercle is preserved proximally. No groove for the flexor tibialis can be observed on this bone as is generally the case in cercopithecines but present in colobines ( Olivier & Fontaine 1957; Strasser & Delson 1987; Gilbert et al. 2010).



| AC |
Amherst College, Beneski Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.