
Sabellomma minuta
|
publication ID |
https://doi.org/10.5281/zenodo.275967 |
|
DOI |
https://doi.org/10.5281/zenodo.6209681 |
|
persistent identifier |
https://treatment.plazi.org/id/03036267-8005-FFFB-FF6B-537BFEB9FF49 |
|
treatment provided by |
Plazi |
|
scientific name |
Sabellomma minuta |
| status |
comb. nov. |
Sabellomma minuta View in CoL comb. nov.
( Figs 1–6 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , 16 View FIGURE 16 A–F; Table 1)
Parasabella minuta Treadwell, 1941: 4 View in CoL , Figs 9–12 View FIGURE 9 View FIGURE 10 View FIGURE 11 View FIGURE 12 .
Perkinsiana minuta View in CoL .- Knight-Jones, 1983: 285 –287, Fig. 19 View FIGURE 19 A–M.
Material examined. Holotype ( AMNH 2894) and 22 specs, all from the northern coast of the state of São Paulo, Brazil. Holotype coll. Ilha de São Sebastião, city of Ilhabela. Ubatuba: Picinguaba, in algae, shallow subtidal: algal frond 2, 8 Oct 2001: spec. 1 ( MZUSP 1045); algal frond 4, 8 Apr 2001: spec. 2 (LACM-AHF POLY 2395); algal frond 5: spec. 3 ( MZUSP 1046); algal frond 6: spec. 4 (ZUEC-POL 7442); algal frond 7: specs 5–7, spec. 5 (LACM-AHF POLY 2396), spec. 6 (LACM-AHF POLY 2397), spec. 7 (ZUEC-POL 7443); algal frond 10: specs 8–11, spec. 8 (LACM-AHF POLY 2398), spec. 9 (LACM-AHF POLY 2399), spec. 10 (LACM-AHF POLY 2400), spec. 11 (discarded); algal frond 11, 8 Jun 2001: spec. 12 (LACM-AHF POLY 2401); algal frond 12: spec. 13 (LACM-AHF POLY 2402); algal frond 17: spec. 14 ( MZUSP 1047); algal frond 19: specs 15–17, spec. 15 ( MZUSP 1048), spec. 16 (ZUEC-POL 7444), spec. 17 ( MZUSP 1049). São Sebastião: Praia de São Francisco, 19 Apr 2003, intertidally, in rocks: specs 18–20, spec. 18 ( MZUSP 1050), spec. 19 (ZUEC-POL 7445), spec. 20 (LACM-AHF POLY 2403); Praia da Baleia, algal frond 4: spec. 21 (ZUEC-POL 7446); islands in front of Praia da Baleia, in Sargassum , shallow subtidal, 10 Apr 2003, fronde 3: spec. 22 ( MZUSP 1051). Details of holotype and specs. 1–22 are provided in Table 1.
Additional material. São Sebastião, Praia de São Francisco, 16 Jul 2003, intertidal, on rocks: 9 specs.
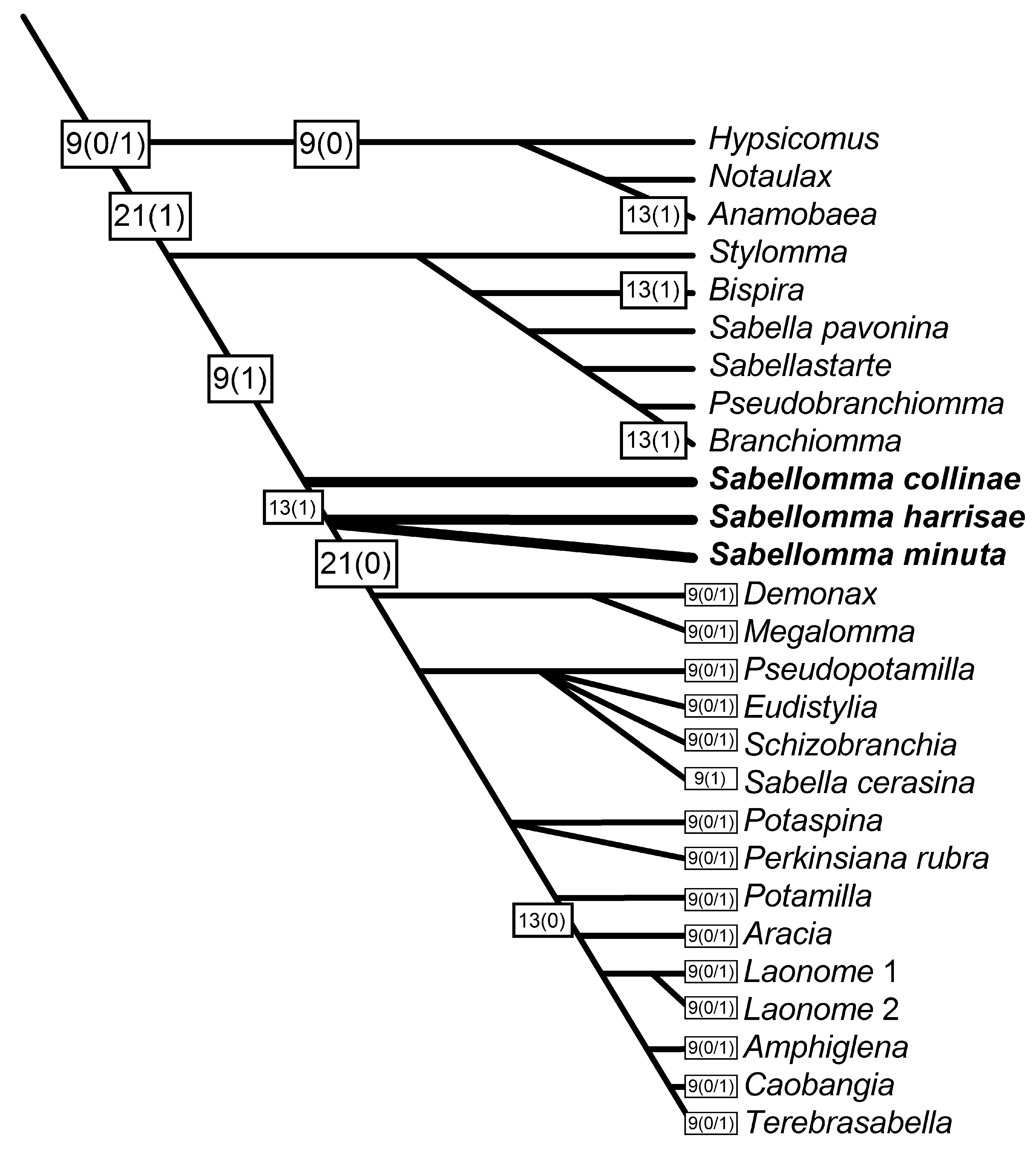
Definition. Two possibilities, depending on character transformation series in the phylogenetic hypothesis involving individuals to which Sabellomma collinae spec. nov., S. minuta comb. nov., and S. harrisae spec. nov. refers (see Remarks on Specific-level Hypotheses in Sabellomma ): (1) A specific hypothesis, accounting for the presence of pinnular appendages fused for almost entire length to dorsal lips among observed individuals. Pinnular appendages almost completely fused to dorsal lips originated by an unspecified mechanism(s) in a reproductively isolated population of individuals with pinnular appendages only proximally fused to dorsal lips, subsequent to which the new character became fixed in the population by an unspecified mechanism(s), leading to individuals observed in the present, all with pinnular appendages almost entirely fused to dorsal lips. (2) There are no autapomorphies to provide a basis for the specific hypothesis.
As is discussed below (see Remarks on Specific-level Hypotheses in Sabellomma ), the formal definition of S. minuta as an explanatory hypothesis is tenuous since the explanation of pinnular appendages completely fused to dorsal lips is not an autapomorphy in some transformation series (see also Definition for S. harrisae , below).
Description. Measurements. Medium-sized individuals; crown 1.7–3.6 mm long, 5–8 pairs of radioles ( Table 1). Complete specimens with trunk 2.5– 9 mm long ( holotype longer but incomplete), 0.3-0.85 mm wide, 3–6 thoracic chaetigers (most with 4–5), 17–52 abdominal chaetigers ( Table 1).
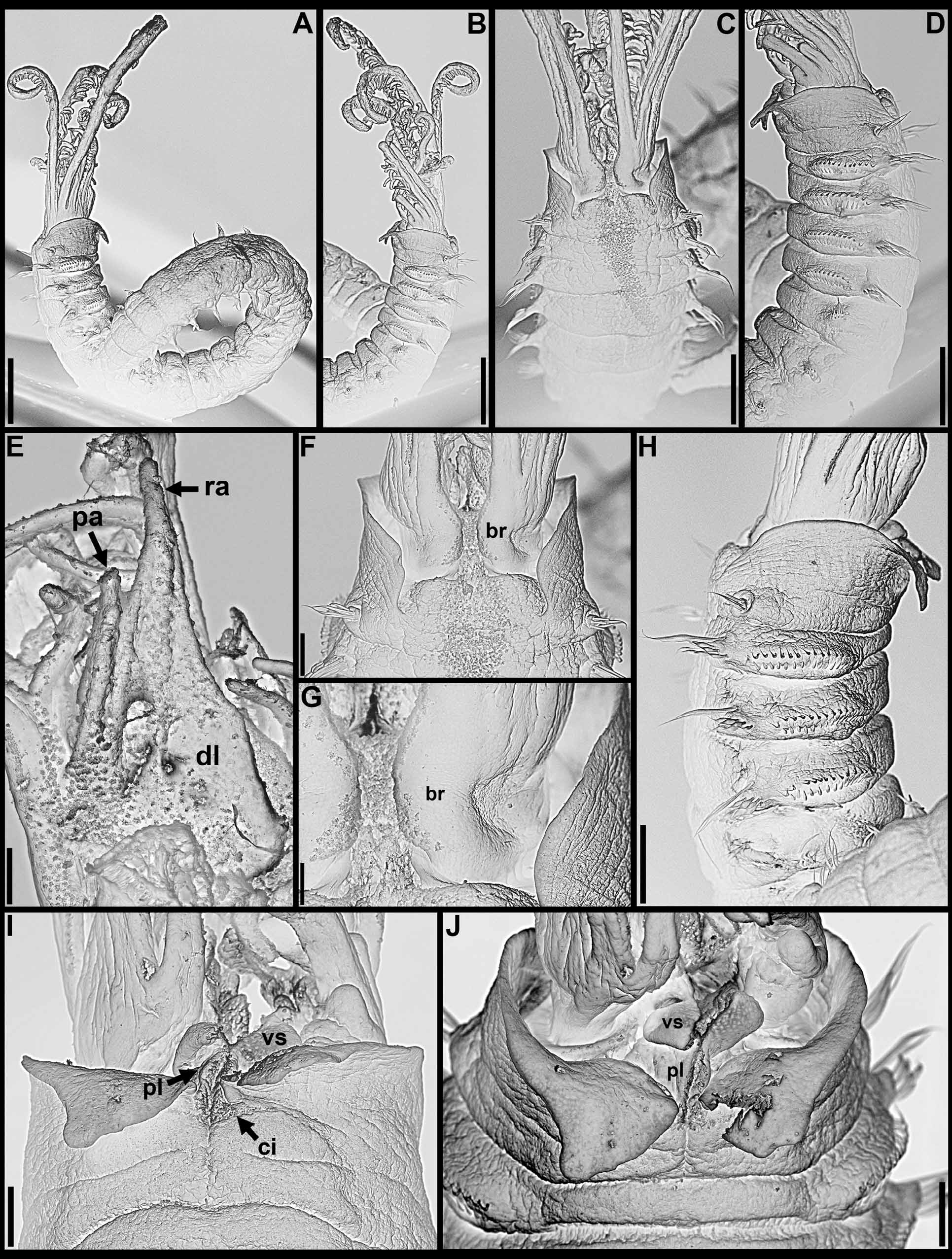
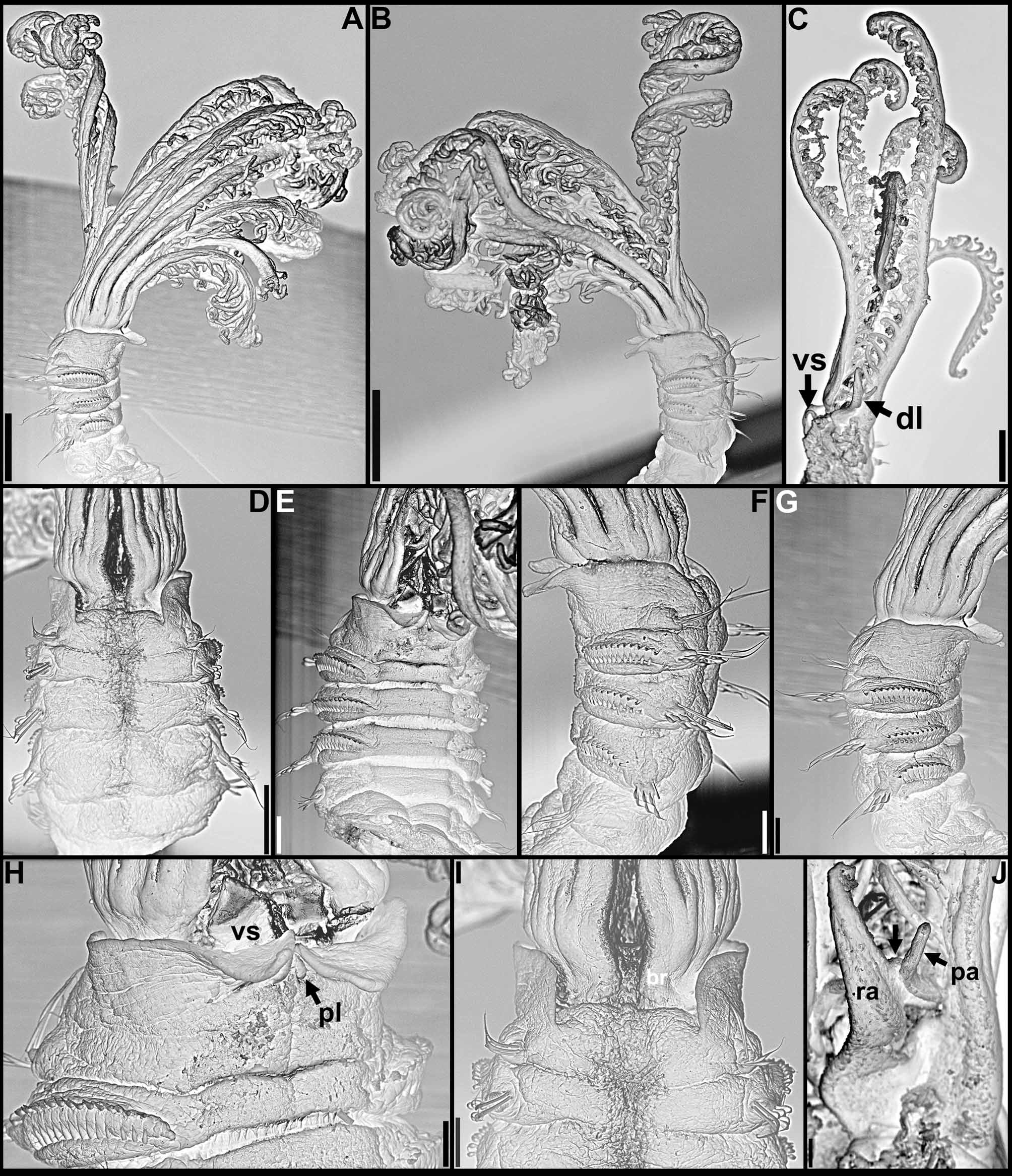
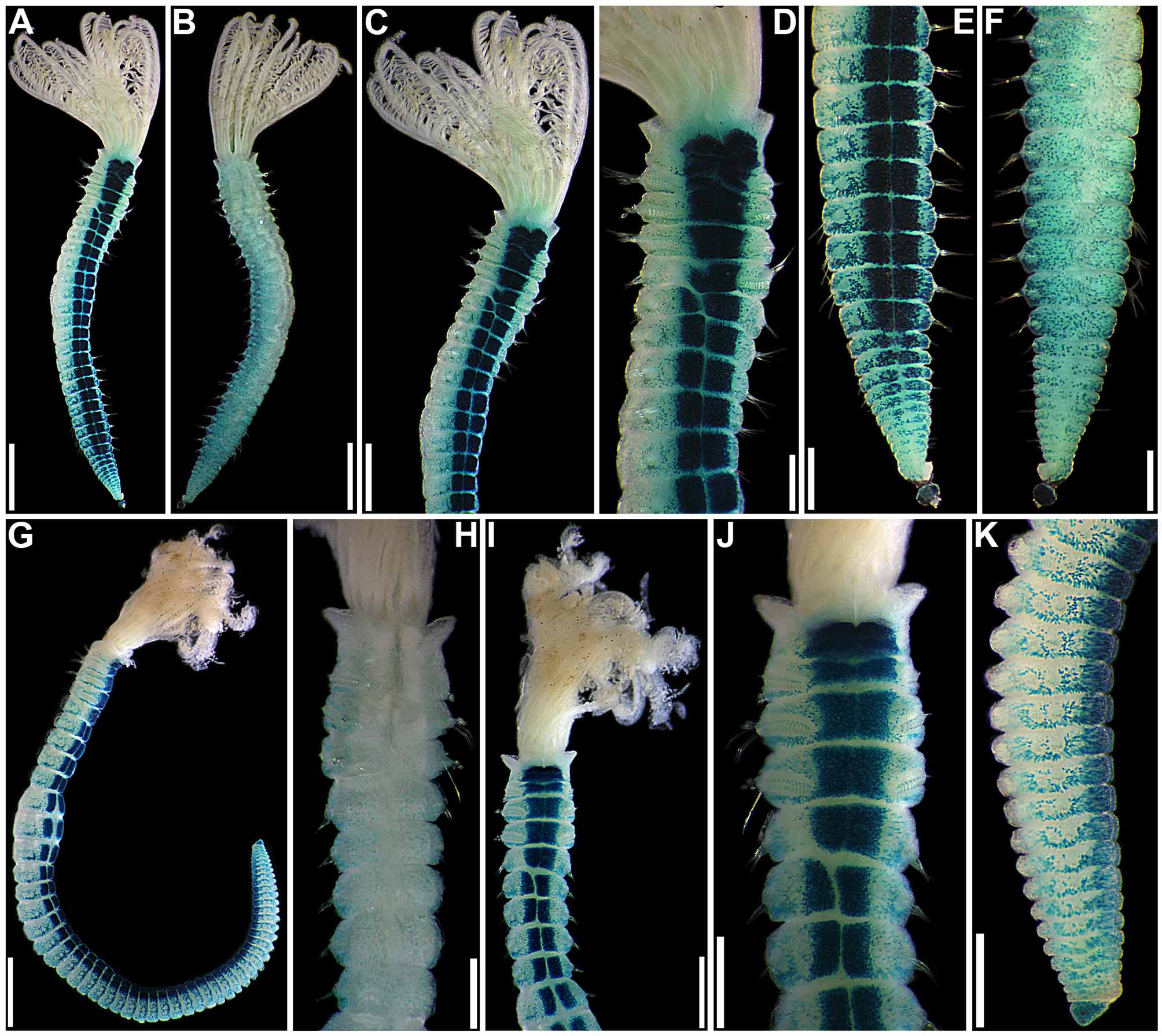
Pigmentation. Live specimens with radioles alternating pink and white bands, associated with dark green and pink pinnules, radiolar tips and associated pinnules white; anterior body brown, small interramal eyespots throughout ( Fig. 1 View FIGURE 1 A). After preservation, anterior body pigmentation and interramal eyespots frequently conspicuous, crown with up to 7 brownish bands, alternating with white bands, pinnules mostly white or with fainted pigmentation ( Fig. 1 View FIGURE 1 B–G).
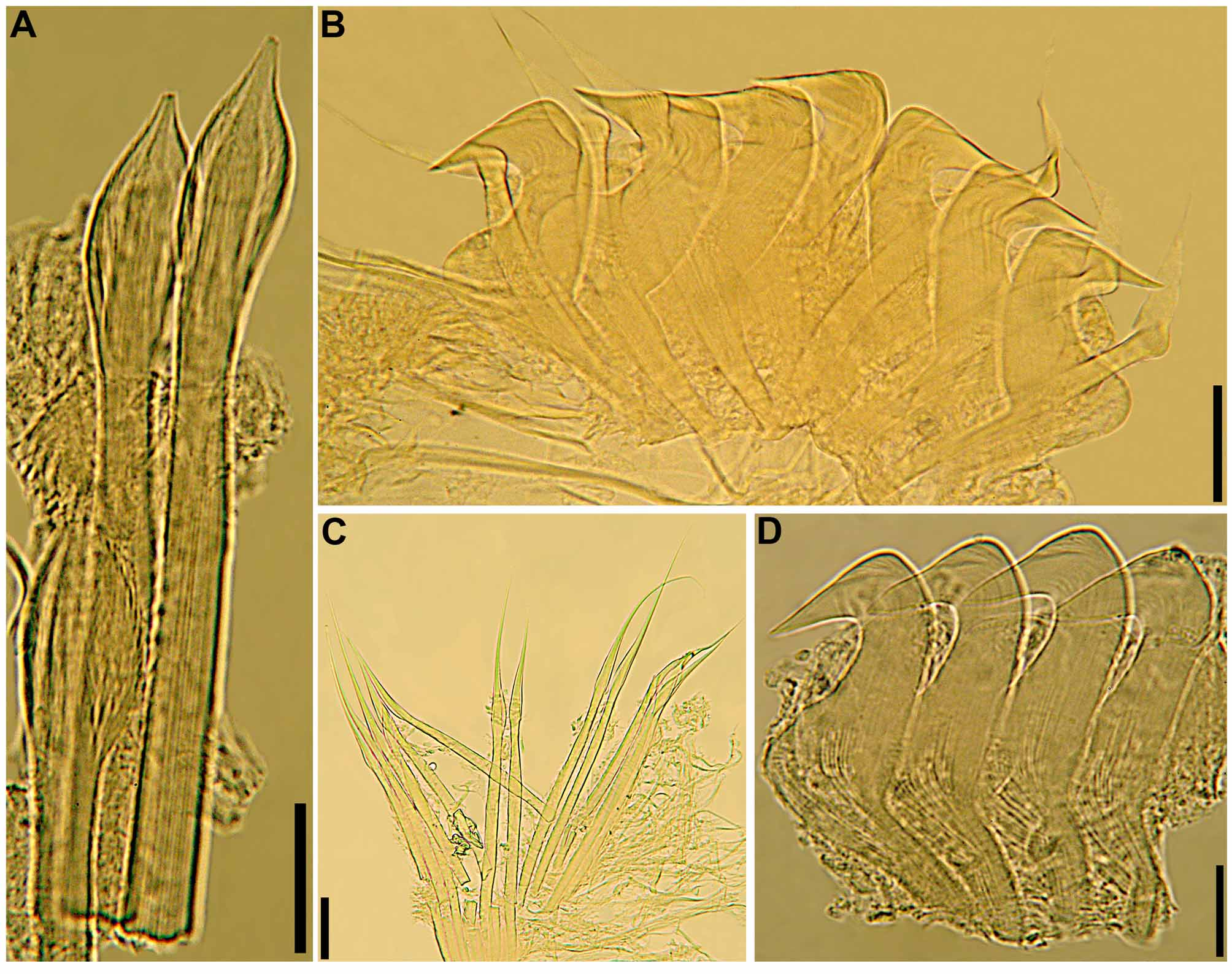
Crown. Radiolar flanges and palmate membrane both absent. Each radiole with four rows of skeletal cells. Radioles with one irregular row of eyespots along each lateral margin; eyespots as single units, each with large lens, more numerous along distal two-thirds to one-half of radioles, especially in association with transverse pigment bands ( Figs 1 View FIGURE 1 F–G; 3A), less abundant near radiolar tips. Dorsal lips about one-fifth total crown length, basally broad, distally tapering, with radiolar appendages lacking internal skeletal cells; single pinnular appendage almost completely joined by thin membrane to each dorsal lip ( Fig. 2 View FIGURE 2 E). Ventral lips low, distally rounded, each continuing ventrally as parallel lamella with large ventral sac above collar ventral lappets ( Figs 1 View FIGURE 1 G; 2B, D, H–J). Mid-dorsal margins of branchial lobes as thickened ridges, extending anteriorly along dorsal-most pair of radioles ( Fig. 2 View FIGURE 2 C, F–G).
Number of segments
Number of pairs
Size of radioles
length of crown; body Thorax (left; Abdomen (left; Slides (segments) Additional data
(left; right sides of
length x width (mm) right sides of right sides of
crown) body) body)
Thorax: notochaetae
(paleae) and neurochaetae: Holotype in relatively good state of preservation, with fainted but Holotype 3 conspicuous radiolar eyespots. Body pigmentation, including
3; 13 (incomplete) x 0.8 8; 7 4 38; 37
AMNH 2894) interramal eyespots, all faded off, not visible. Photographed for Figs
Abdomen: neurochaetae 1B, E; 6A–D
and uncini: 9
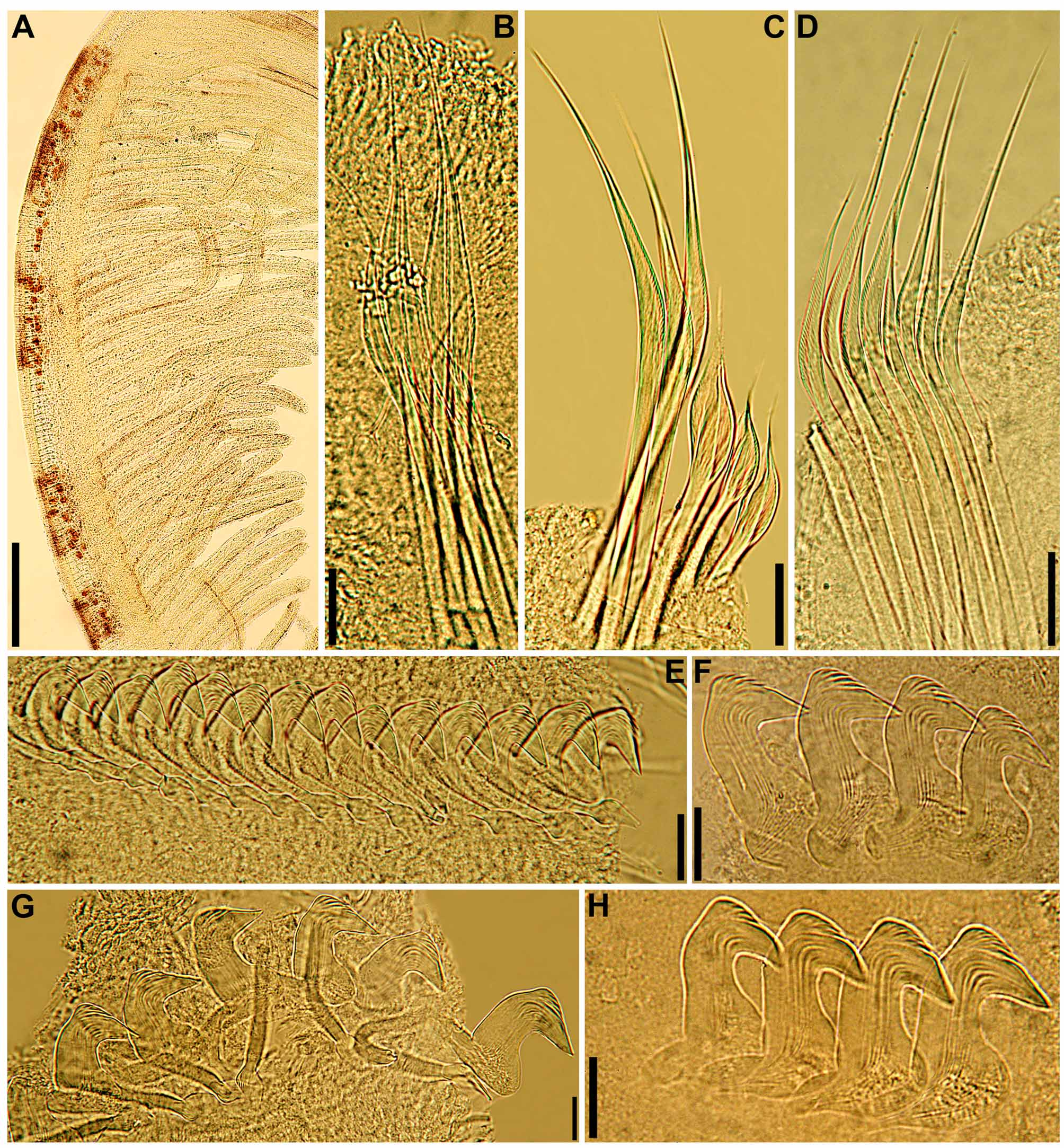
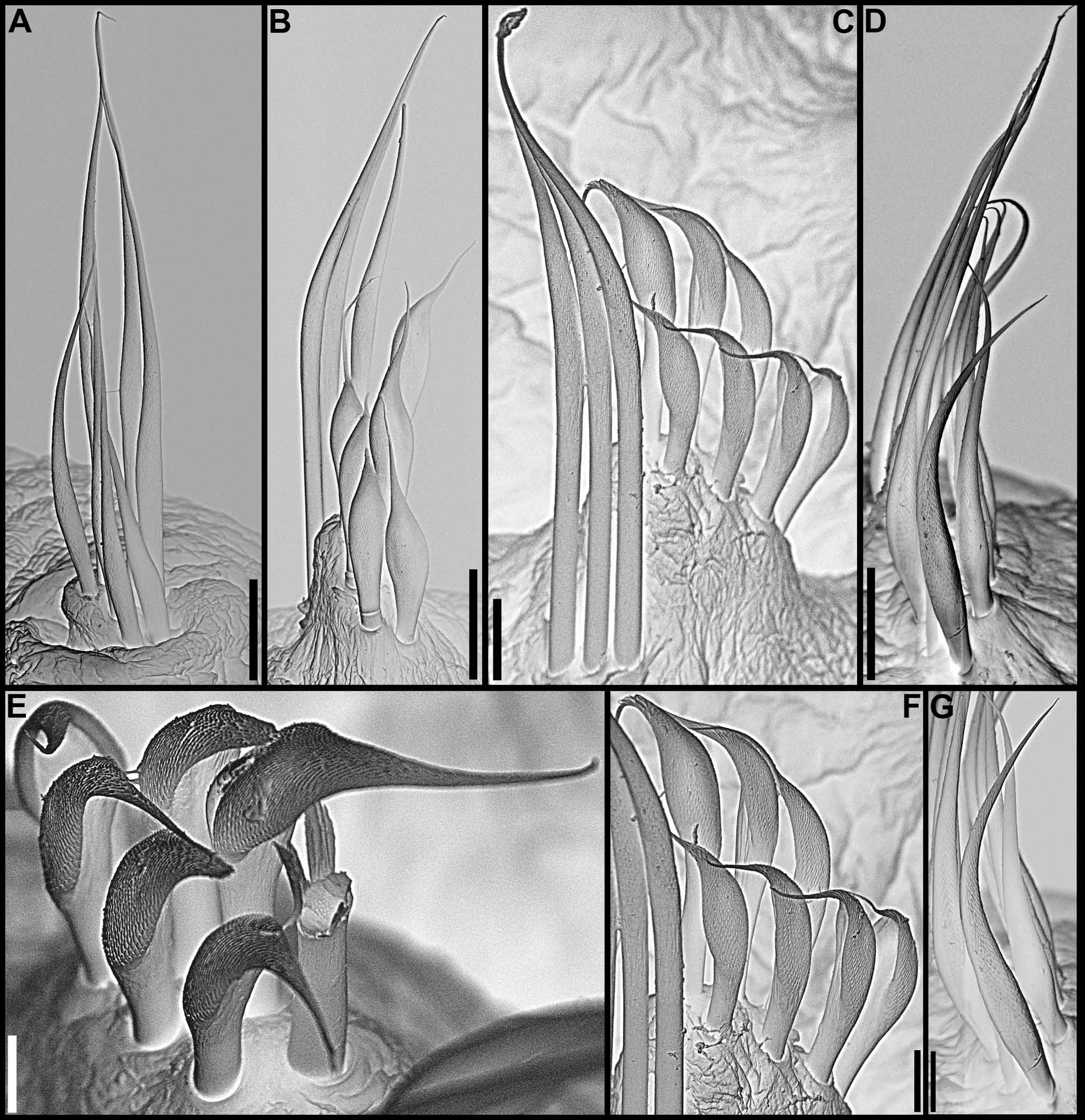
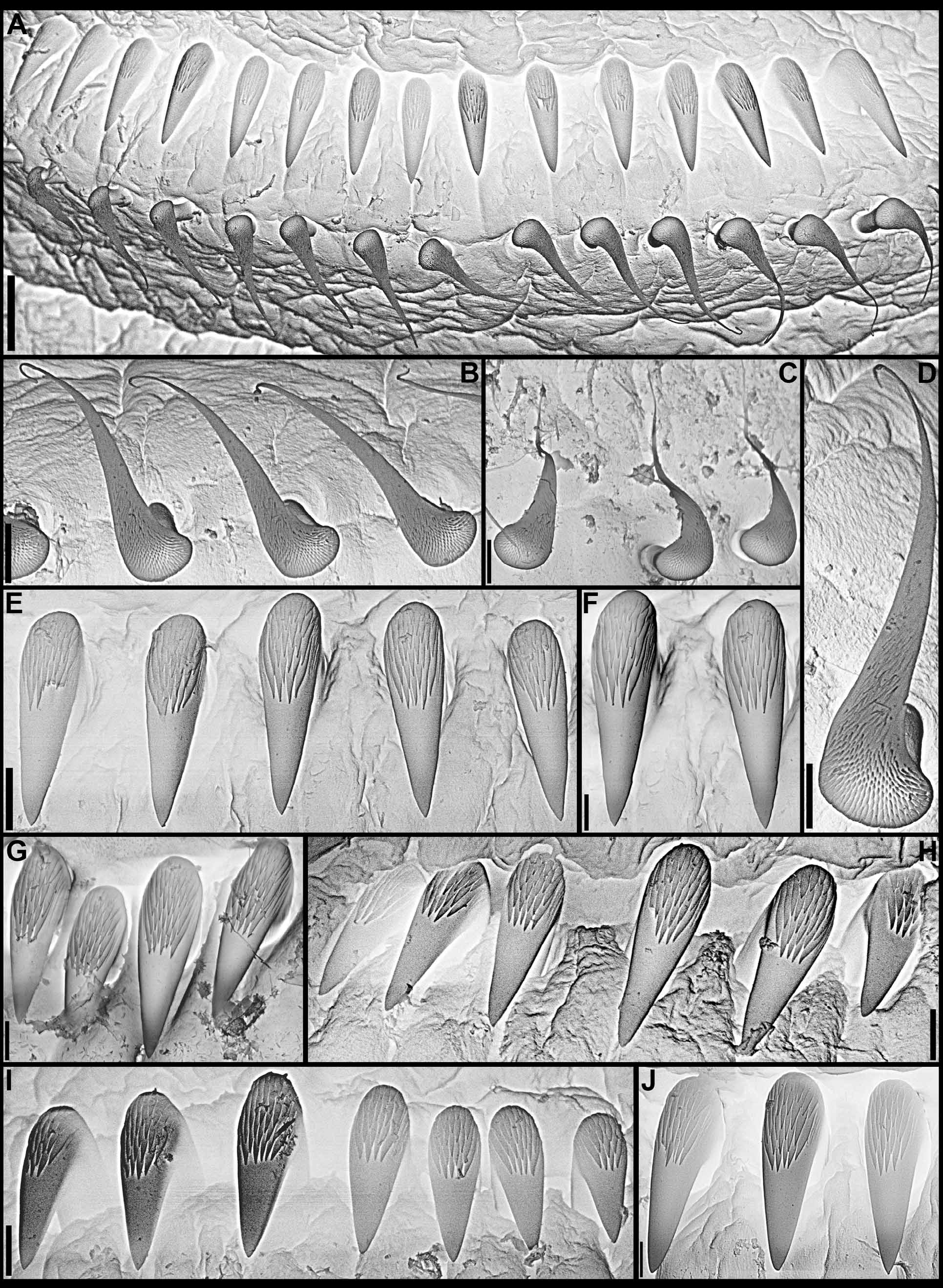
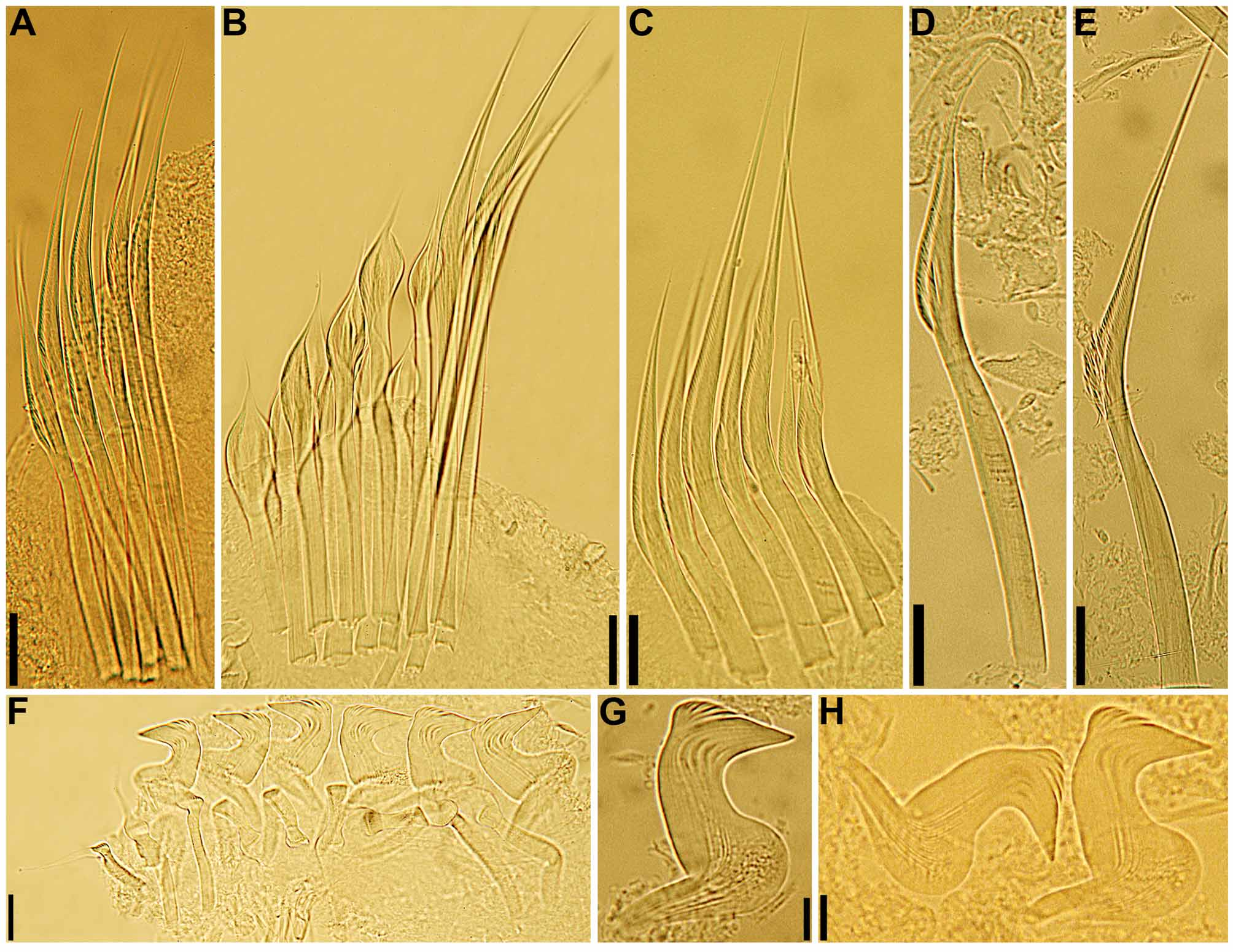
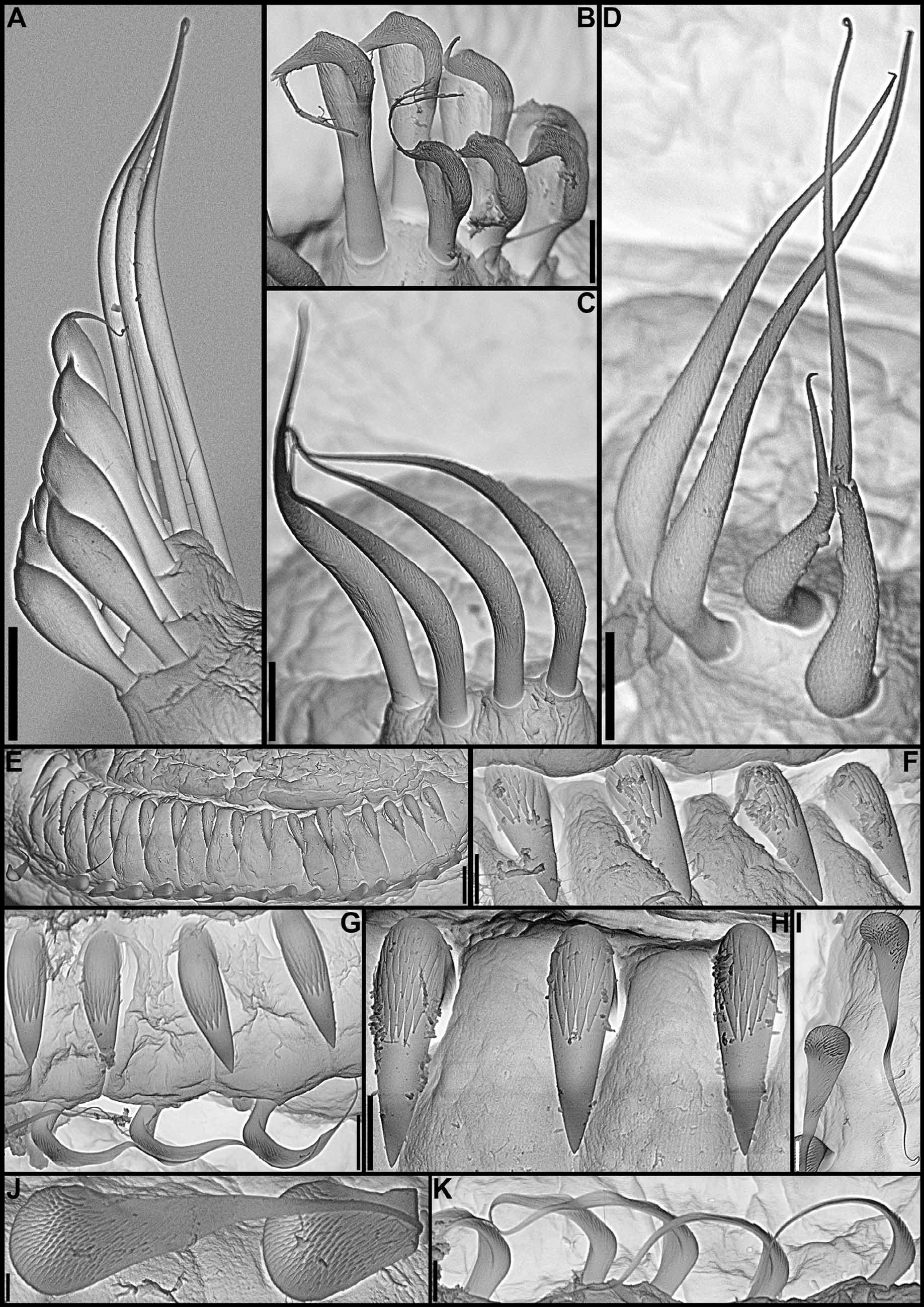
continued next page Body. Collar fused to body dorso-laterally, well separated from faecal groove, extending anteriorly to level of first pair of parapodia; laterally low, slightly oblique, covering bases of radiolar crown but not origins of radioles; ventrally with one pair of triangular, distally pointed, non-overlapping lappets reaching anterior margin of chaetiger 1, separated from each other by short parallel lamellae; ventrally, between termination of parallel lamellae and anterior margin of first ventral shield, ciliated patch present ( Figs 1 View FIGURE 1 B–E, G; 2A–D, F–J). Trapezoidal ventral shields, indented by neuropodia on thorax; anterior margin of first shield medially indented ( Figs 1 View FIGURE 1 B–C, E–G; 2B, D, H). Mid-dorsal faecal groove deep on anterior chaetigers ( Fig. 2 View FIGURE 2 C, F). Collar chaetae narrowly hooded, arranged in two rows ( Figs 2 View FIGURE 2 A–D, F, H, J; 3B, 4A); subsequent thoracic notopodia with narrowly hooded superior chaetae, arranged in arc, inferior chaetae paleate, with long, gently tapering tips, arranged in two rows partially enclosed by superior arc ( Figs 2 View FIGURE 2 A–D, H, J; 3C; 4B–C, E–F). Thoracic neuropodia with avicular uncini about as long as high, crest with 4–5 irregular rows of secondary teeth, covering proximal half of main fang, breast distinct, not extending distally to tip of main fang ( Figs 3 View FIGURE 3 E, G; 5A, E–F); companion chaetae with rounded, symmetrical, denticulate heads and long, tear drop-shaped, slightly asymmetrical, gently tapering membranes ( Figs 3 View FIGURE 3 E; 5A–D). Abdominal neuropodia with narrowly hooded chaetae, arranged in two rows, anterior row with shorter chaetae ( Figs 3 View FIGURE 3 D; 4D, G). Anterior abdominal uncini similar to thoracic ( Figs 3 View FIGURE 3 F; 5G), from mid-abdomen uncini with similar number of rows of secondary teeth ( Fig. 5 View FIGURE 5 H–J), but with shorter handles ( Fig. 3 View FIGURE 3 H). Triangular, distally rounded pygidium, with 1–3 small, red ocelli at each lateral margin ( Figs 1 View FIGURE 1 H, 16K).
Remarks on holotype. The holotype is relatively well preserved, but most pigmentation of the crown and body is faded and inconspicuous, including that of interramal eyespots, except for conspicuous radiolar eyespots on most radioles. Radiolar eyespots were noticed by both Treadwell (1941) and Knight-Jones (1983), although in her diagnosis of Perkinsiana, Knight-Jones (1983) stated that radiolar eyes are absent among members of the genus.
Knight-Jones (1983) examined the holotype and made slides of a thoracic and an abdominal chaetiger. All data about the holotype provided by Knight-Jones is accurate, except for some misinterpretations.
Thoracic notopodial paleae ( Fig. 6 View FIGURE 6 A), and abdominal neuro- ( Fig. 6 View FIGURE 6 C) and notochaetae ( Fig. 6 View FIGURE 6 D) of the holotype are like those of recently collected specimens. Although Knight-Jones (1983) stated that abdominal neurochaetae are similar in length throughout, they do vary along the body; the holotype has longer chaetae in posterior rows starting in mid-abdominal segments. The only real discrepancy between the redescription provided by Knight-Jones (1983) and in this paper is in regards to the lengths of handles in thoracic uncini, which, according to Knight-Jones’ drawing (1983: 286, Fig. 19 View FIGURE 19 G), is about twice the length of distance between breast and crest. However, the slide of thoracic neurochaetae of the holotype made by Knight-Jones has the torus of segment 3 split into three pieces and in two of them the shafts of companion chaetae completely cover the handles of uncini, not allowing for proper evaluation of their length, and in the third piece only a few uncini have visible handles, which are clearly much shorter than the shafts of companion chaetae ( Fig. 6 View FIGURE 6 B).
The holotype is also much longer than any of the other specimens examined in the present study ( Table 1), but the presence of a short thorax and radiolar eyespots, together with the fact that it was collected on São Sebastião Island, which is across a 2 km wide channel from the beaches where our specimens were collected, give us confidence that the hypothesis Sabellomma minuta applies to all of the specimens. In addition, the area has been extensively surveyed for more than five years and the only other Sabellinae specimens found with a short thorax are members of Pseudobranchiomma paraemersoni Nogueira, Rossi & López, 2006 .
Variation. The specimens of Sabellomma minuta examined for the present study presented great variation in size and crown/body pigmentation ( Table 1). Since these conditions have not yet been compared to what exists in live specimens, it is not possible to discount the effects of preservation.
The range of variation in numbers of thoracic chaetigers is 3–6 among observed specimens ( Table 1). This variation might reflect the consequences of regeneration subsequent to scissiparity ( Knight-Jones & Bowden 1984; Fitzhugh 2003; Nogueira & Knight-Jones 2003; Nogueira et al. 2004), especially considering the amount of variation between left and half sides of the body in number of radioles and body segments, which we believe is also due to imperfect regeneration.
Distribution. This species is only known to occur off the northern coast of the State of São Paulo, Brazil. Previous records by Knight-Jones (1983) from the Caribbean are herein assigned to S. collinae spec. nov., (see below).


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Sabellinae |
|
Genus |
Sabellomma minuta
| Nogueira, João Miguel De Matos, Fitzhugh, Kirk & Rossi, Maíra Cappellani Silva 2010 |
Perkinsiana minuta
| Knight-Jones 1983: 285 |
Parasabella minuta
| Treadwell 1941: 4 |