
RHINOCRYPTIDAE, WETMORE, 1926
|
publication ID |
https://doi.org/10.1111/j.1096-3642.2012.00847.x |
|
persistent identifier |
https://treatment.plazi.org/id/03800613-174E-FFFF-271A-FCDFFB83721A |
|
treatment provided by |
Marcus |
|
scientific name |
RHINOCRYPTIDAE |
| status |
|
MONOPHYLY OF RHINOCRYPTIDAE View in CoL
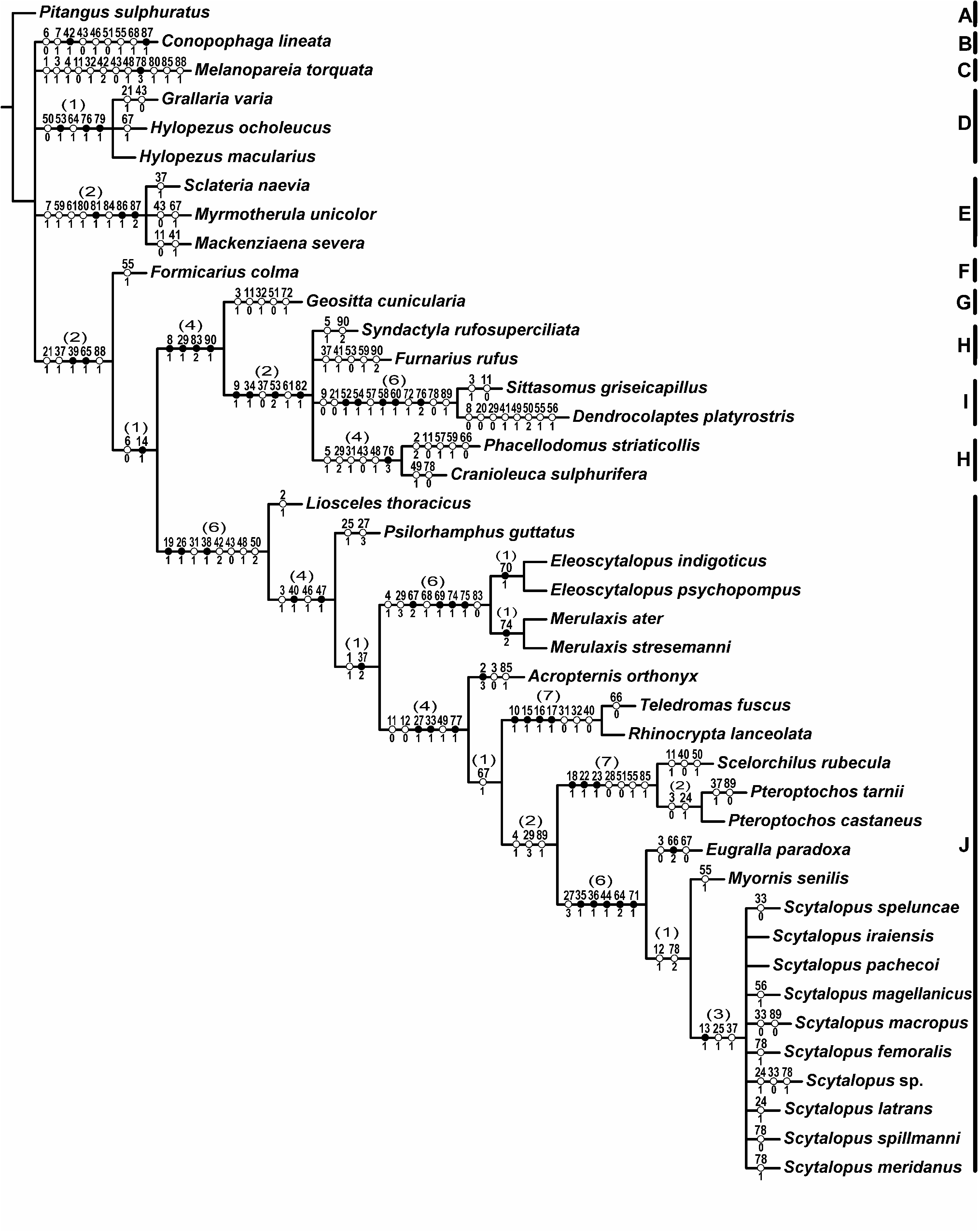
The present morphology-based analysis constitutes a robust test of monophyly of the family as it includes representatives of all families of the infraorder Furnariides and all genera of Rhinocryptidae – including a heterogeneous sampling of the highly speciose genus Scytalopus . The only other study aimed at testing the monophyly of the family was the recent sequence-based analysis presented by Ericson et al. (2010), which included representatives of most families of Furnariides (except Scleruridae) and all rhinocryptid genera, but a much less heterogeneous sampling of Scytalopus . The two studies are therefore complementary. The Rhinocryptidae as recovered in our morphological phylogeny is perfectly congruent with all molecular phylogenies with adequate taxonomic coverage ( Moyle et al., 2009; Ericson et al., 2010) in excluding the genus Melanopareia from the family and supporting the allocation of the problematic genera Psilorhamphus and Teledromas within Rhinocryptidae . The resulting monophyletic Rhinocryptidae has strong Bremer support (i.e. 6), includes 12 genera ( Liosceles , Psilorhamphus , Eleoscytalopus , Merulaxis , Acropternis , Rhinocrypta , Teledromas , Pteroptochos , Scelorchilus , Eugralla , Myornis , and Scytalopus ), and is defined by the following eight synapomorphies, the first four of which are features not previously mentioned for the family: nonpneumatized ectethmoid (char. 19.1), absence of foramen in the medial face of the brachial tuberosity of the coracoid (char. 43.0), ulna as long as or shorter than humerus (char. 48.1), distance between the posterior projections of the ilium nearly equal to the width of the caudalmost sacral vertebrae (char. 50.2), absence of a foramen in the medial process of the mandible (char. 38.1), caudal margin of the sternum with two pairs of notches (char. 42.2), lacrimal partially fused to the ectethmoid (char. 26.1), and parietal and frontal bones largely non-pneumatized (char. 31.1). Additionally, the last four characters have never been formally suggested as synapomorphic for the family, although the last two have been regarded as ‘characteristics that unite members of the tapaculo family’ (termed ‘lachrymal bones fused into the ectethmoid’ and ‘incomplete skull ossification’, respectively) by Krabbe & Schulenberg (2003: 752). The four-notched sternum was optimized as a synapomorphy of the Rhinocryptidae in the only previous morphology-based cladistic study that included members of the family ( Pteroptochos and Rhinocrypta ) ( Claramunt & Rinderknecht, 2005), judging from the consensus tree and the data matrix presented.
Below, a detailed account regarding the systematic placement of each of the ‘problematic’ genera is provided.
Melanopareia: Comprehensive View in CoL taxon and character sampling is particularly important to elucidate the position of Melanopareia View in CoL because morphology-based studies ( Ames, 1971; Feduccia & Olson, 1982) found it natural to place Melanopareia View in CoL within Rhinocryptidae View in CoL , as first suggested by W. W. Miller on the basis of this genus’ possession of a four-notched sternum ( Wetmore, 1926). However, these morphological studies did not include cladistic analyses of characters and based their decisions solely upon particular features whose systematic value is subjective if not analysed within a strict cladistic context. Feduccia & Olson (1982) noted that Melanopareia View in CoL lacked some features typical of rhinocryptids such as the absence of a foramen in the medial process of the mandible but concluded that ‘... our examination showed its osteology to be in accordance with its placement in the Rhinocryptidae’ (p. 6). However, apart from some general similarities, one can find only two objective characters in the literature that would support Melanopareia View in CoL as part of the Rhinocryptidae View in CoL , namely the possession of a lacrimal partially fused to the ectethmoid and the four-notched sternum ( Feduccia & Olson, 1982; Krabbe & Schulenberg, 2003; Ericson et al., 2010). On the basis of material available for the present study, however, the lacrimal was not partially fused to the ectethmoid in Melanopareia View in CoL , and instead its lacrimal was found to be completely fused to the ectethmoid with no signs of suture, as in the great majority of the members of Furnariides . On the other hand, a number of character-states – including the four-notched sternum – were found here to occur in the Rhinocryptidae and View in CoL in the Melanopareiidae View in CoL ( Fig. 43 View Figure 43 ), but these shared conditions were solidly optimized in the parsimony analysis as convergences between distantly related groups: Melanopareia View in CoL was part of a polytomy with the families Conopophagidae View in CoL , Grallariidae View in CoL , Thamnophilidae View in CoL , and a large, structured clade that included Formicariidae View in CoL , Scleruridae, Furnariidae View in CoL , Dendrocolaptidae , and Rhinocryptidae View in CoL . Another morphological feature that supports the exclusion of Melanopareia View in CoL from the Rhinocryptidae View in CoL is its retention of the primitive morphology of the stapes (flat footplate) instead of the typical suboscine condition present in all rhinocryptids (inflated footplate), including Psilorhamphus View in CoL and Teledromas View in CoL ( Feduccia, 1974; Ericson et al., 2010). In any event, the present morphology-based phylogeny corroborates the erection of the family-level taxon Melanopareiidae View in CoL for the genus Melanopareia View in CoL as first proposed by Irestedt et al. (2002) and later formalized by Ericson et al. (2010).
Psilorhamphus: Although View in CoL it has been suggested that Psilorhamphus View in CoL might lie outside Rhinocryptidae ( Ridgely & Tudor, 1994) View in CoL and that it bears little external resemblance to typical members of the family ( Krabbe & Schulenberg, 2003; Ericson et al., 2010), Psilorhamphus View in CoL proved to be a typical member of the group in terms of osteology, with 12 synapomorphies supporting it as a rhinocryptid (eight in the family node and four in the subsequent node) and no character state linking it to other family of Furnariides (see Fig. 43 View Figure 43 ). Therefore, the allocation of the genus to the Rhinocryptidae View in CoL as first implemented by Plótnick (1958) was a correct systematic decision, with two of the characters that supported this author’s decision –M. sternotrachealis originating in the vertebral segment of the second rib and presence of two pairs of notches in the sternum – having been cladistically optimized here as support for such a placement (the first character for the Formicariidae View in CoL + Furnariidae View in CoL + Scleruridae + Dendrocolaptidae + Rhinocryptidae View in CoL clade and the latter for the rhinocryptid clade). Recent molecular phylogenies also corroborated this placement ( Maurício et al., 2008; Ericson et al., 2010).
Teledromas: Based View in CoL on similarity of vocalizations ( Ridgely & Tudor, 1994; N. Krabbe, unpubl. data), general shape, details of the nasal operculum, tarsal scutellation, flank pterylography, and straight shape of humerus between Teledromas View in CoL and Melanopareia, Irestedt et al. (2002) View in CoL suggested that they were members of the same clade. Given that Melanopareia View in CoL proved to be a distinct lineage of uncertain affinities, they placed both genera in the new family Melanopareiidae View in CoL even though no sequence data of Teledromas View in CoL were available. Subsequent molecular phylogenies that included Teledromas View in CoL samples showed this genus to be well embedded within Rhinocryptidae View in CoL ( Moyle et al., 2009; Ericson et al., 2010), a placement corroborated in the present analysis. In fact, anatomically Teledromas View in CoL is very similar to Rhinocrypta View in CoL , both having several shared derived character states (see above). The alleged similarity of Teledromas View in CoL to Melanopareia ( Irestedt et al., 2002) View in CoL , at least in terms of general aspect and shape of the humerus, found no support here. Moreover, although vocalizations were considered as the main evidence for a close link between Teledromas View in CoL and Melanopareia View in CoL ( Irestedt et al., 2002; Ericson et al., 2010) detailed structural analyses of the vocal repertoire of Teledromas View in CoL support its inclusion within the Rhinocryptidae View in CoL (J. I. Areta, unpubl. data).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
RHINOCRYPTIDAE
| Maurício, Giovanni Nachtigall, Areta, Juan Ignacio, Bornschein, Marcos Ricardo & Reis, Roberto E. 2012 |
Melanopareia
| Irestedt 2002 |
Rhinocryptidae
| WETMORE 1926 |
Rhinocryptidae
| WETMORE 1926 |
Teledromas
| Wetmore and Peters 1922 |
Rhinocrypta
| G. R. Gray 1841 |
Formicariidae
| G. R. Gray 1840 |
Formicariidae
| G. R. Gray 1840 |
Thamnophilidae
| Swainson 1824 |