
Austrachelas wassenaari, Haddad, Charles R., Lyle, Robin, Bosselaers, Jan & Ramirez, Martin J., 2009
|
publication ID |
https://doi.org/10.5281/zenodo.191594 |
|
DOI |
https://doi.org/10.5281/zenodo.6225471 |
|
persistent identifier |
https://treatment.plazi.org/id/038087A2-DA58-4501-FF4F-FA366127FAFF |
|
treatment provided by |
Plazi |
|
scientific name |
Austrachelas wassenaari |
| status |
sp. nov. |
Austrachelas wassenaari View in CoL n. sp.
( Figs 14, 15 View FIGURES 1 – 15 , 78–81 View FIGURES 76 – 81 )
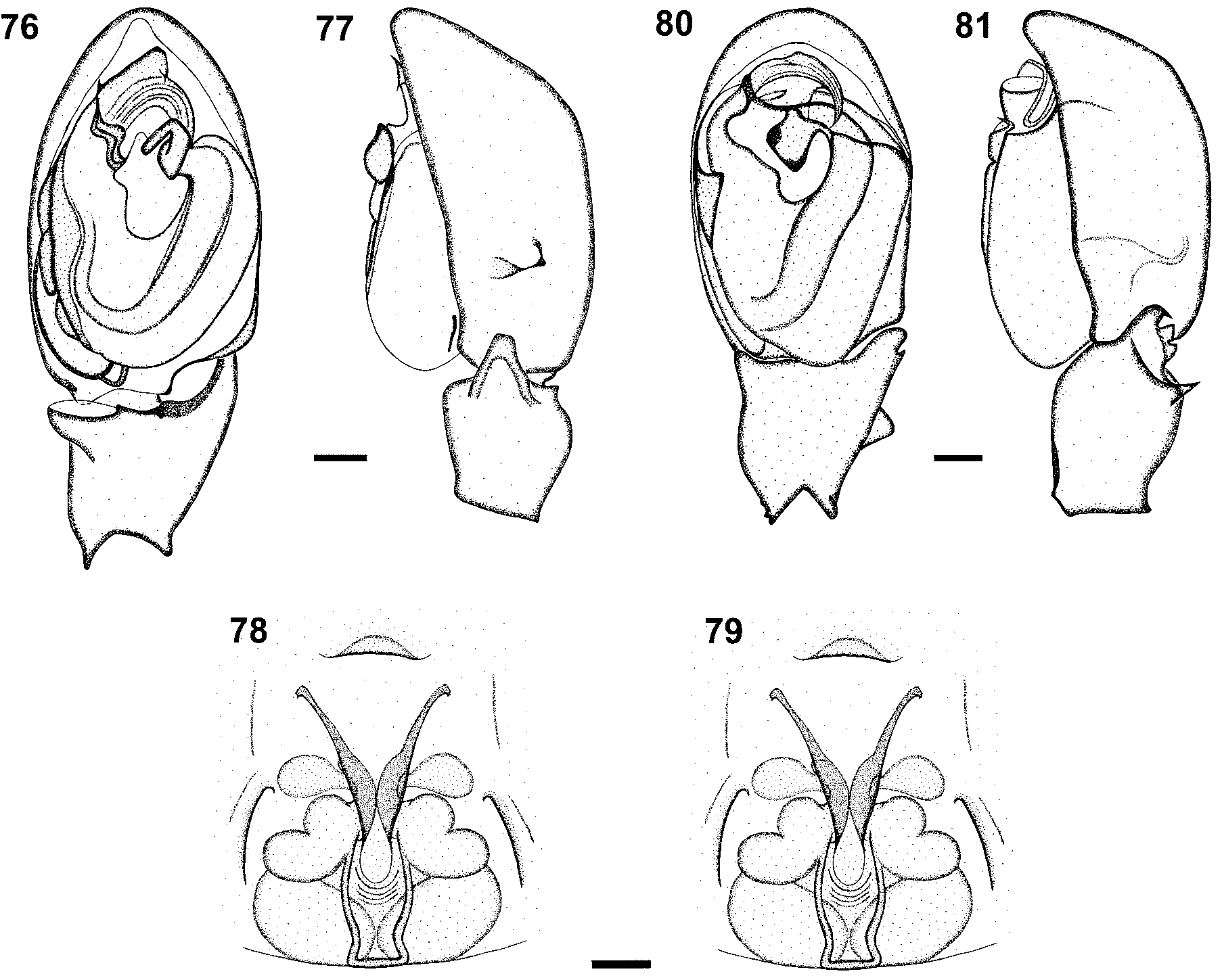
Diagnosis. This species is related to A. natalensis and females can be distinguished by the larger spermathecal structures and narrower anterior hood of the epigyne ( Fig. 78 View FIGURES 76 – 81 ), and males by the structure of the retrolateral tibial apophysis of the palp, with a sharp dorsal tooth ( Fig. 81 View FIGURES 76 – 81 ).
Etymology. This species is named after Theo Wassenaar, who collected many of the type specimens.
Female. Measurements: CL 2.87–3.10, CW 2.03–2.25, FL 0.28–0.38, AL 3.35–4.20, AW 1.63–2.18, SL 1.60–1.70, SW 1.18–1.28, TL 6.25–7.40, AME–AME 0.05, AME–ALE 0.03, ALE–ALE 0.28, PME–PME 0.05, PME–PLE 0.10, PLE–PLE 0.45.
Length of leg segments (sequence from femur to tarsus, and total): I 2.08 + 1.35 + 1.55 + 1.05 + 0.78 = 6.81; II 1.95 + 1.20 + 1.43 + 1.05 + 0.68 = 6.31; III 1.95 + 0.80 + 0.93 + 1.28 + 0.75 = 5.71; IV 2.35 + 1.20 + 1.78 + 2.15 + 0.88 = 8.36.
Carapace deep orange-brown, orange along midline ( Fig. 14 View FIGURES 1 – 15 ); AER strongly procurved, laterals larger than medians; AME separated by distance equal to ½ their diameter, AME separated from ALE by distance equal to ½ AME diameter; clypeus height equal to 1¼ AME diameter at AME, equal to ½ ALE diameter at ALE; PER very slightly recurved, nearly straight, laterals larger than medians; PME separated by distance equal to ½ their diameter, PME separated from PLE by distance equal to PME diameter. Chelicerae deep orange; promargin with four evenly spaced teeth, proximal tooth smallest, second tooth largest, third and distal teeth intermediate in size, subequal in size; retromargin with single small denticle, situated near third promarginal tooth. Legs all yellow-orange in colour, paler distally. Leg spination: femora: I pl 1 do 1, II pl 1 do 1, III pl 2 do 2 rl 1, IV do 2; patellae: spineless; tibiae: I & II spineless, III pl 3 rl 2 plv 2 rlv 1 vt 2, IV pl 3 rl 2 plv 2 rlv 2 vt 2; metatarsi: I & II spineless, strongly scopulate, III & IV scopulate in distal half, III pl 6 rl 4 plv 1 rlv 1 vt 2, IV pl 6 rl 5 plv 2–3 rlv 1 vt 2; palp: femora do 2, patellae spineless, with prominent pl 1 do 2 setae, tibiae pl 4 rl 2, tarsus pl 3 rl 3 plv 1 rlv 1. Abdomen dark grey dorsally, with mottled pale grey chevron ( Fig. 14 View FIGURES 1 – 15 ), paler grey laterally and ventrally; dorsum and venter lacking scutum or other sclerites. Female epigyne with distinctive lateral hoods and narrow anterior hood ( Fig. 78 View FIGURES 76 – 81 ); copulatory openings anterolaterally situated, entrance ducts narrow, entering broad C-shaped ST 2 with anterolateral lobe, connected closely to large posteriorly situated oval ST 1 ( Fig. 79 View FIGURES 76 – 81 ).
Male. Measurements: CL 3.35–3.63, CW 2.48–2.83, FL 0.37–0.48, AL 2.95–3.40, AW 1.83–2.30, SL 1.80–2.00, SW 1.35–1.58, TL 6.15–7.30, AME–AME 0.05, AME–ALE 0.04, ALE–ALE 0.30, PME–PME 0.07, PME–PLE 0.11, PLE–PLE 0.51.
Length of leg segments (sequence from femur to tarsus, and total): I 2.60 + 1.45 + 2.00 + 1.45 + 0.98 = 8.48; II 2.48 + 1.43 + 1.70 + 1.45 + 0.95 = 7.66; III 2.10 + 1.13 + 1.25 + 1.65 + 0.85 = 6.98; IV 2.83 + 1.40 + 2.15 + 2.68 + 1.10 = 10.16.
Carapace deep wine-red, black around margins, paler along midline ( Fig. 15 View FIGURES 1 – 15 ); AER strongly procurved, laterals slightly larger than medians; AME separated by distance equal to ½ their diameter, separated from ALE by distance equal to ¼ AME diameter; clypeus height equal to 1¼ AME diameter at AME, equal to ¾ ALE diameter at ALE; PER very slightly recurved, nearly straight, laterals larger than medians; PME separated by distance equal to ½ their diameter, PME separated from PLE by distance equal to PME diameter. Chelicerae deep red-brown, anterior margin with coarse ridges; promargin with two teeth, distal tooth largest and proximal tooth smallest; retromargin with two teeth, smaller than promarginal teeth, situated closer to fang base. Legs all deep orange-brown, paler orange distally. Leg spination: femora: I pl 1 do 1, II pl 1–2 do 2, III pl 2–7 do 3 rl 1, IV do 2; patellae: spineless; tibiae: I & II spineless, III pl 3 rl 2 plv 2 rlv 1 vt 2, IV pl 2 rl 2–3 plv 2 rlv 2 vt 2; metatarsi: I & II spineless, strongly scopulate, III & IV scopulate in distal half, III pl 6 rl 4–6 plv 1 rlv 1 vt 2, IV pl 6 rl 4–5 plv 2 rlv 1 vt 2. Abdomen dark grey with pale grey chevron dorsally ( Fig. 15 View FIGURES 1 – 15 ), cream laterally and ventrally; dorsum with small anterior scutum, venter without sclerites. Palpal median tegular apophysis broad, hook-like; embolus arc-shaped, distal margin of embolus without lobes or teeth ( Fig. 80 View FIGURES 76 – 81 ); palpal tibia with hook-like retrolateral apophysis with two closely associated pointed lobes and sharp dorsal tooth-like apophysis ( Fig. 81 View FIGURES 76 – 81 ); palpal spination: femora do 2.
Type material: Holotype Ƥ: SOUTH AFRICA: KwaZulu-Natal Province, Richards Bay, [ 28°43'S, 32°12'E], 26.VIII.1996, T. Wassenaar (pit traps) (NCA 97/110).
Paratypes: SOUTH AFRICA: KwaZulu-Natal Province, Richard’s Bay, [ 28°43'S, 32°12'E], 18.VI.1996, T. Wassenaar (pitfall traps), 13 2Ƥ (NCA 97/72); Same data, 29.VII.1996, 23 1Ƥ (NCA 97/73); Greater St. Lucia Wetlands Park, Fanies Island, 28°32'S, 32°24'E, 21-25.VII.1990, M. Alderweireldt & R. Jocqué (pitfall, woodland edge with grass), 13 2Ƥ ( MRAC 171752).
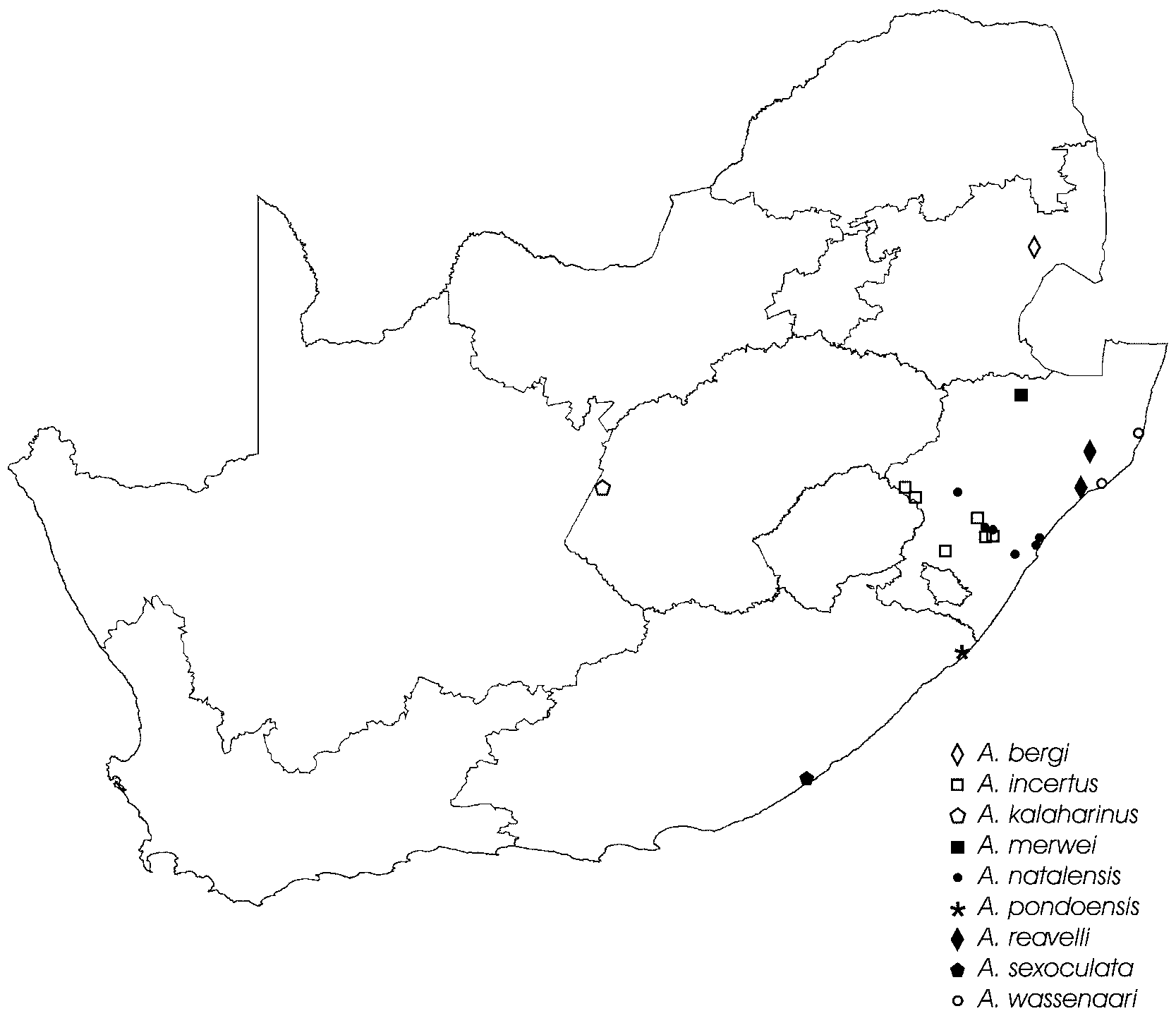
Distribution. Known from northern coastal parts of KwaZulu-Natal, South Africa ( Fig. 82 View FIGURE 82 ).
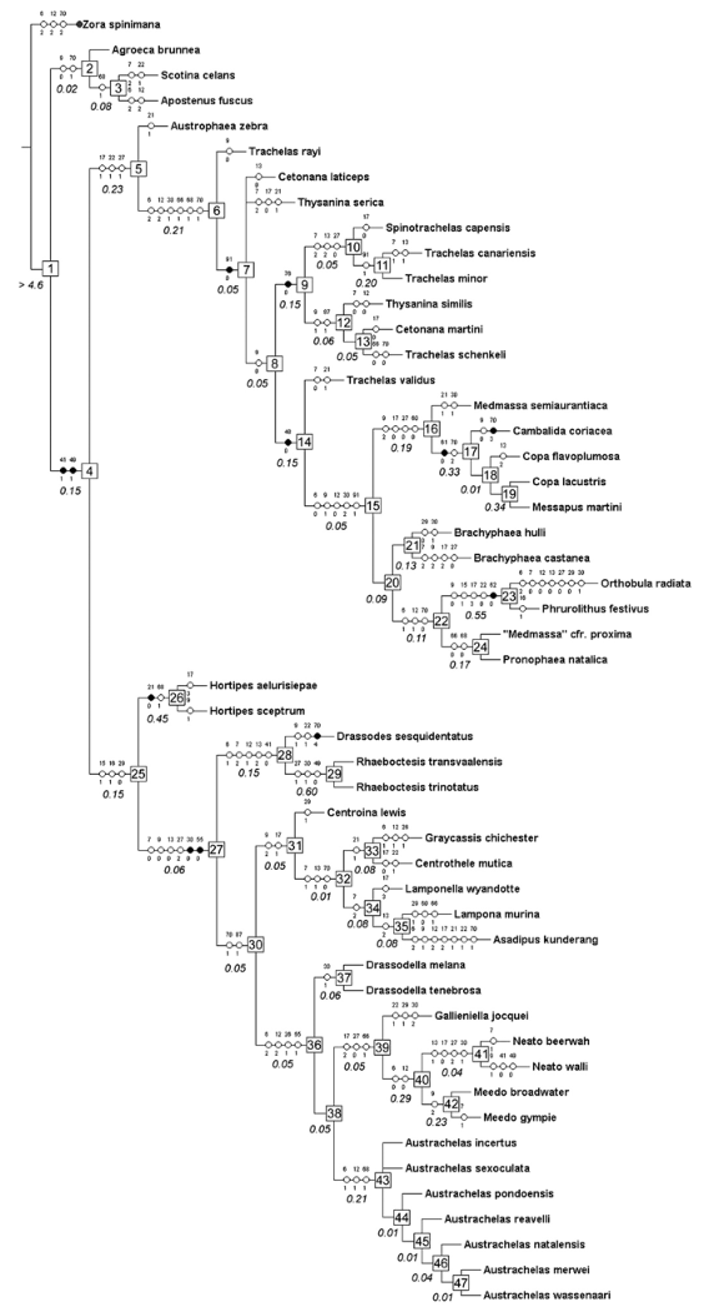
The genus Austrachelas now comprises nine species endemic to South Africa. The species generally occur in moist savanna and forest habitats in the east and south-east of the country, with the exception of A. kalaharinus n. sp., which occurs in dry Acacia savanna in central South Africa. Consistent with other Gallieniellidae , Austrachelas are ground-dwelling spiders that were captured mainly by pitfall trapping.
The somatic morphology of the genus is very stable, reflected in the results of the phylogenetic analysis ( Fig. 50 View FIGURE 50 , Appendix 2). Male palpal morphology is also very conservative, and species can be separated primarily on small differences in the shape and position of the palpal median apophysis, embolic protuberances and shape of the retrolateral tibial apophysis. Females show more variation than males, particularly in the shape and position of the anterior and lateral epigynal hoods and spermathecal structures.
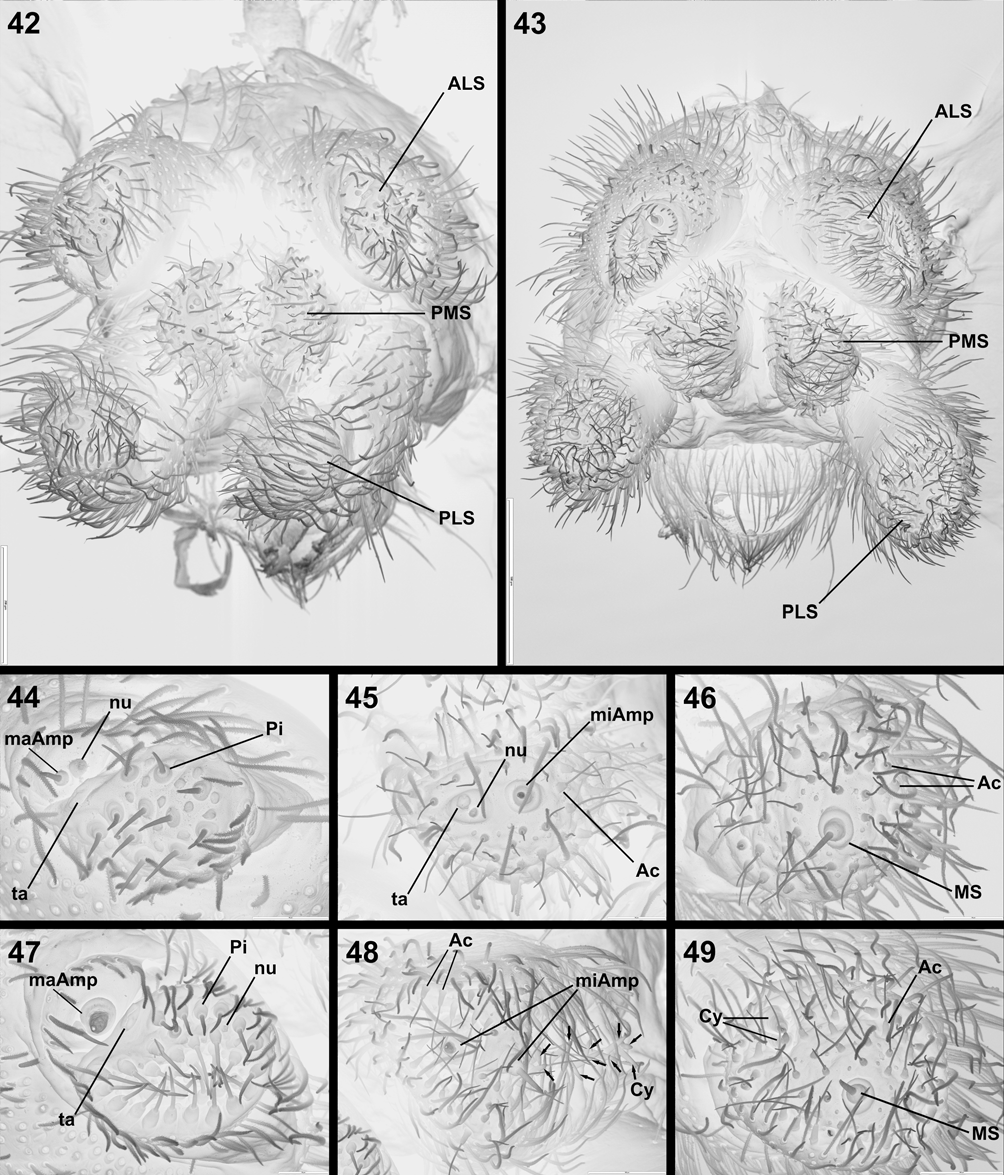
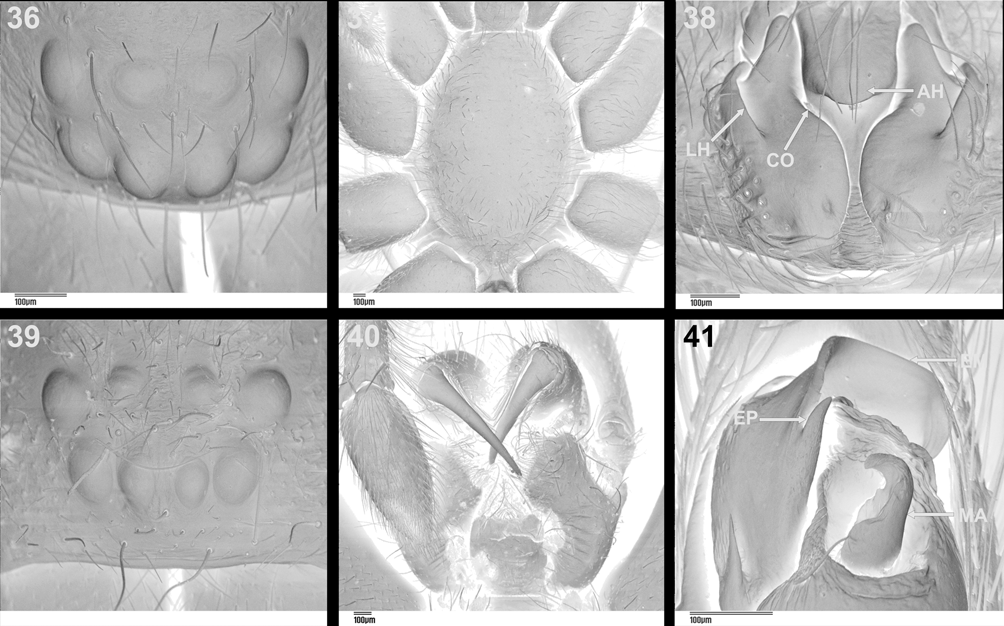
The phylogenetic analysis confirmed earlier suspicions ( Lyle & Haddad 2006b) that Austrachelas is misplaced and should be transferred to Gallieniellidae . Several synapomorphic characters supporting its placement within the Gnaphosoidea include the depressed endites, oval flattened PME and the sclerotised ALS ( Platnick 1984). Further, the “lower gnaphosoid” families have a small distal segment on the ALS, which is at least partly sclerotised ( Platnick 2002), also found in Austrachelas ( Figs 44, 47 View FIGURES 42 – 49 ). The presence of two parallel rows of cylindrical gland spigots on the female PMS of Austrachelas ( Fig. 48 View FIGURES 42 – 49 ) is a characteristic shared by Gallieniellidae and Trochanteriidae within the “lower gnaphosoids” ( Platnick 2002). In addition to the above, its placement within the Gallieniellidae is supported by the following synapomorphies: PME flattened, oval and smaller than PLE ( Figs 36, 39 View FIGURES 36 – 41 ), leg IV longer than leg I, and chelicerae with long, oblique fangs ( Platnick 1984, 2002).
The relationships of Austrachelas within the Gallieniellidae are not yet fully resolved as not all of the known genera were included in our phylogenetic analysis. The genus may be the sister group of Drassodella + Toxoniella , which were suggested by Warui & Jocqué (2002) as possible sister taxa. Austrachelas shares with Drassodella the anterior and lateral epigynal hoods, the hook-like median apophysis on the male palp, the slightly angled chelicerae, and the short dorsal abdominal scutum in males. It differs from Drassodella in the eye arrangement (PER straight rather than slightly recurved), the densely scopulate anterior legs, the chevron-patterned abdomen (as opposed to stripes or spots), oval rather than shield-shaped sternum, and the modified RTA of the male palp (simple and triangular in Drassodella ). Despite these morphological similarities, Drassodella branched as sister to a gallieniellid clade including the type genus Gallieniella , the Australian genera Meedo and Neato , and Austrachelas ( Fig. 50 View FIGURE 50 ). Clearly further studies are necessary to evaluate the phylogeny of the family as a whole.
The present study increases the generic complement of Gallieniellidae in the Afrotropical region to five, and the total family species richness worldwide beyond 50 species (Table 2). Recent taxonomic studies have reported the family for the first time from South America ( Goloboff 2000) and Australia ( Platnick 2002). Thus, the family has a distribution known mainly from the southern hemisphere across all continents.

| MRAC |
Musée Royal de l’Afrique Centrale |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |