
Eunicolina glaesi, Bertrand & Sidorchuk & Hoffeins, 2015
|
publication ID |
https://doi.org/ 10.1051/acarologia/20152170 |
|
persistent identifier |
https://treatment.plazi.org/id/0381FC5B-C662-FFF4-AB92-3FD8FE5BF871 |
|
treatment provided by |
Marcus |
|
scientific name |
Eunicolina glaesi |
| status |
sp. nov. |
Eunicolina glaesi n. sp.
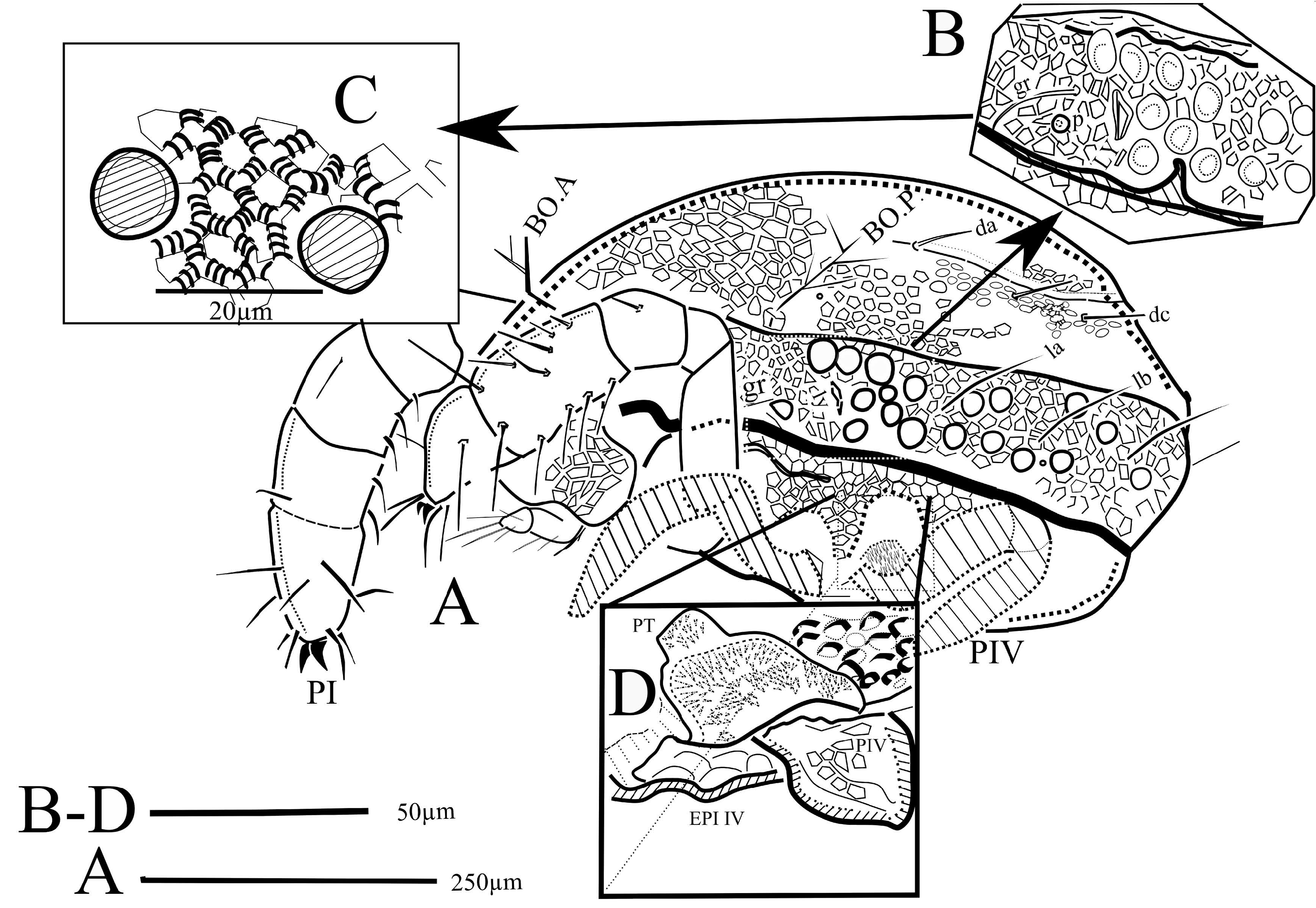
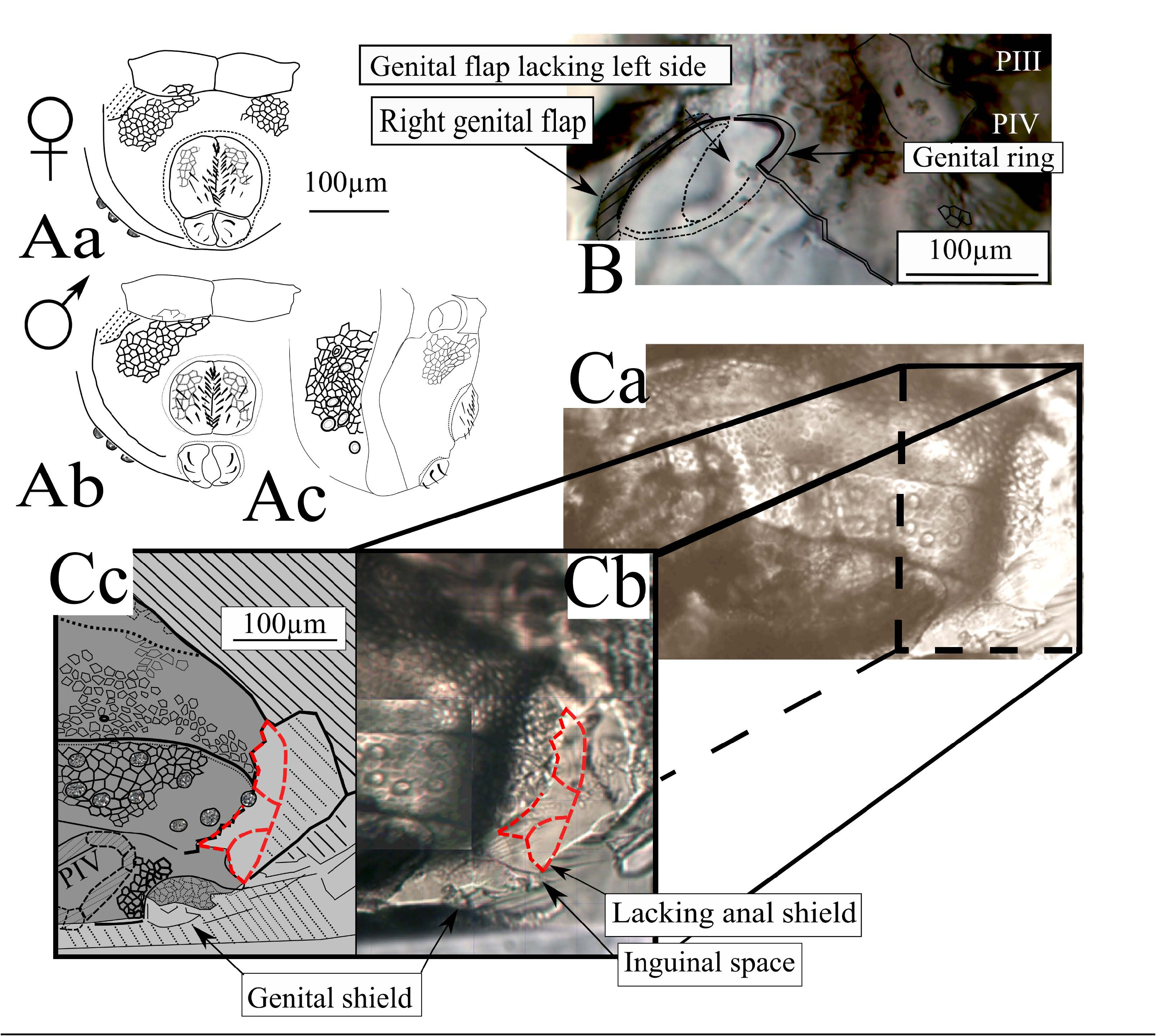
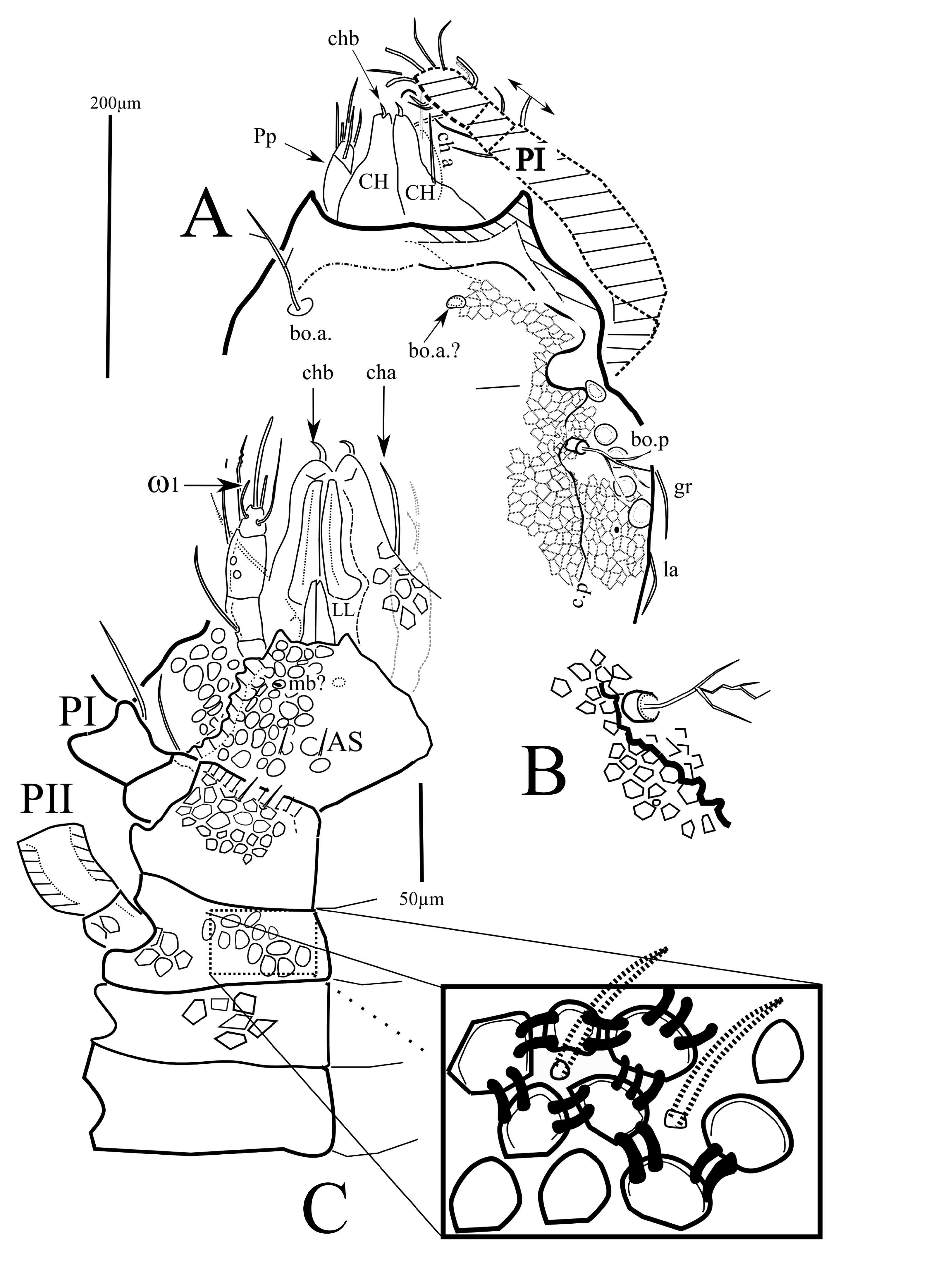
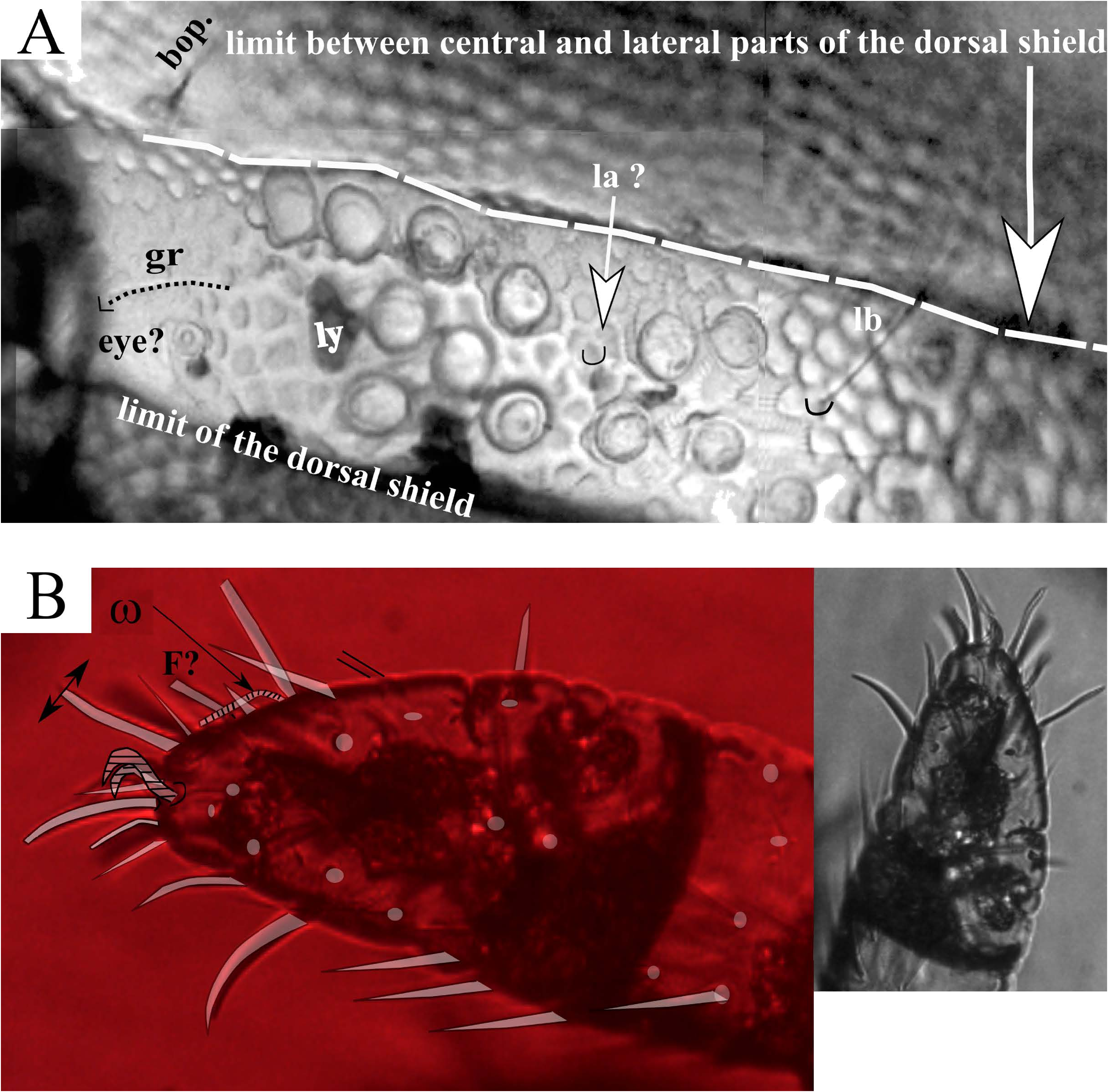
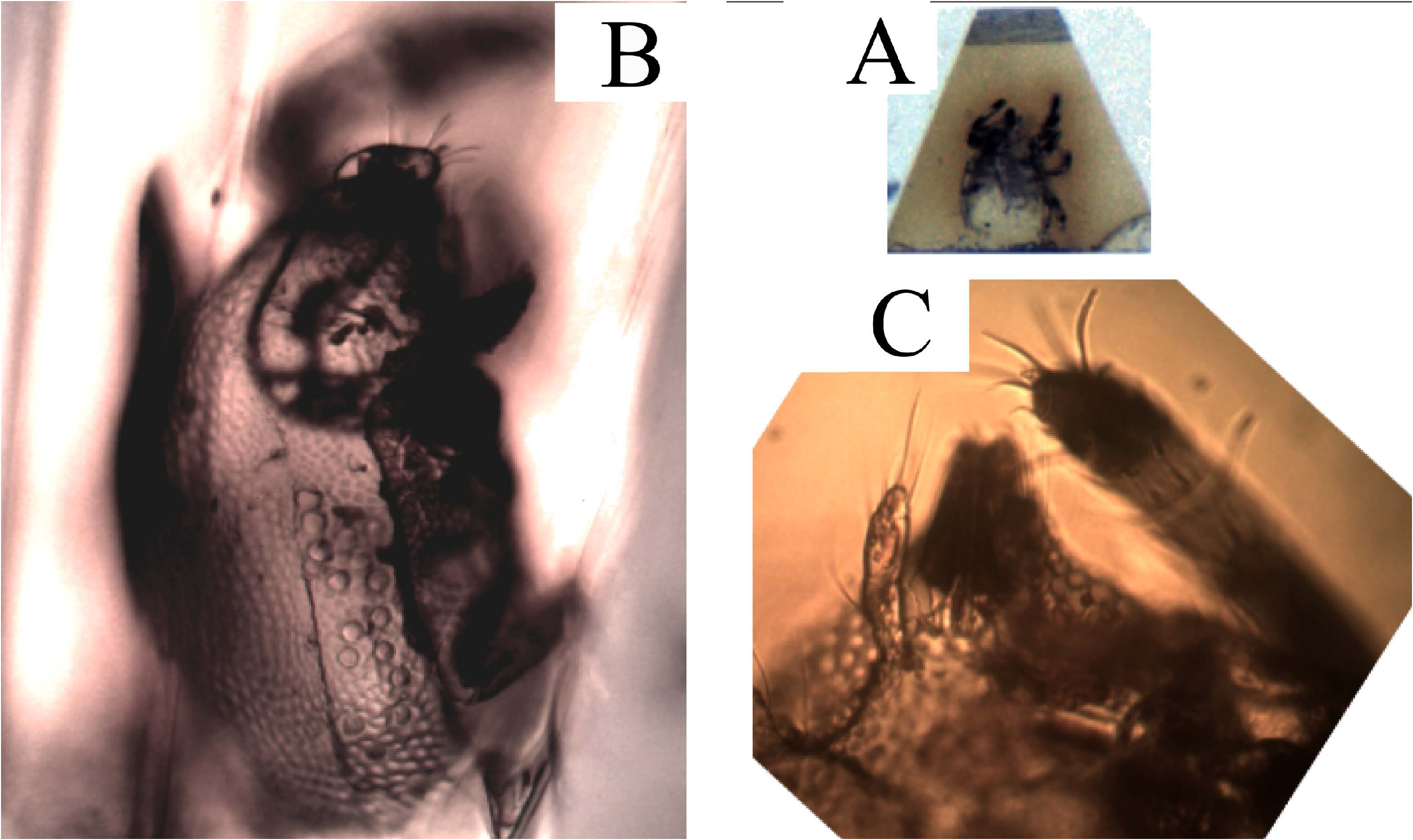
( Figs 1-5 View FIGURE , 6B, 6C View FIGURE )
Type designation and repository — The holotype and only described specimen is an inclusion in Baltic amber from the private C. & H.W. Hoffeins Collection (specimen CHHC 588-13 , which will be eventually transferred to the Museum für Naturkunde, Berlin). This specimen is an adult, having the entire dorsal shield (nymphal stases of labidostommatids are less chitinized, with the dorsal shield divided into paired lateral and dorsolateral sclerotized zones), and, having the rounded genital ring ( Fig. 6 View FIGURE ), it is a male (labidostommatid females have an oval ano-genital ring) .
Description — The habitus is that of an animal with a rounded body, robust, well-sclerotized, yellow to brown in coloration.
Dimensions and proportions — Body length (c. 580 – 600 µm) is about 1.7 times its maximum width (c. 390 – 400 µm), about as wide as high ( Figs 1A View FIGURE , 3B, 3C View FIGURE ).
Cuticle ornamentation and cerotegument — The cuticle is ornamented with more or less rounded alveoli: the dorsal shield is covered with a regular polygonal pattern laterally, whereas partitions are more rounded on the central zone of the dorsum, and on the ventral shields. The borders of alveoli are crossed with more or less large costules ( Fig. 1C View FIGURE ). As in other representatives of the genus, the polygonal pattern extends onto the articles of the legs and on the chelicerae, but not on the palps.
Dorsal shield — Chaetotaxy as for the genus ( Figs 1A View FIGURE , 3A View FIGURE ), the dorsal setae are long, (la – le c. 75 – 100 µm, da – de c. 75 – 80 µm), anterior setae ga, ge, and gm less visible or broken, their presence revealed only by the mark of insertion. The setae gr are short (<40 µm). The trichobothria are similar to those of E. travei Coineau, 1964 , with 3-4 branches. With more than 100 µm in length, the bothridial setae are longer than the lateral and the dorsal setae ( Figs 3A, B View FIGURE ). The frontal edge of the dorsal shield bears two cornua. Close to twenty pustules were counted on the lateral fields of the dorsal shield. On each side, the field of the pustules begins at the level of the second coxal plate. In the usual position of the lateral eyes, close to the (gr) setae, a convex structure ( Figs 1A, 1B View FIGURE , star) could be either a pustule or the lateral eye (the usual striation of the lens surface, different from the smooth surface of the pustules, is not visible). No median anterior eye was discernible. The lateral lyrifissure, poorly preserved, may be located between the most anterior pustules. The density of the pustules is maximal behind the supposed lateral eye ( Figs 1A, 1B View FIGURE , 5A View FIGURE ).
Ventral view — The posterior end of the ventral shield is damaged, hence the anal sclerites were not seen and the genital ones were only partially observable. The infracapitulum (162 µm long, 150 µm wide) is ornamented by alveoli, the pair of setae mb is well visible, plus at least one pair of additional posterior setae ( AS on Fig. 3C View FIGURE ). The lateral lips are narrower than the anterior margin of the infracapitulum. The palps (the right palp was well observable in ventral view) are characteristic for the genus (1-1-3-4+ ω), with the palpal solenidion identifiable and the long apical seta (eupathidium?) (45 – 60 µm long) ( Figs 3C View FIGURE & 4C View FIGURE ). The chelicerae are partially hidden by the first pair of legs and the palps, and cannot be observed in lateral view. The chelicerae are robust (c. 250 µm long) and are strongly sclerotized, the proximal part covered with alveolar ornamentation. Examination in alternate dorsal and ventral views permitted identification of the anterior seta (chb) of the fixed digit and the long posterior seta cha, whose insertion was not seen ( Figs 3A, 3C View FIGURE ). The coxisternal area ( Fig. 3C View FIGURE ) is armoured, sclerotized and ornamented with strong alveoli ( Fig. 3C View FIGURE ), the epimeral setae are poorly preserved, and only some simple setae were identified on the anterior fringe of the first epimeral plates.
At the level of the fourth epimera, on each side, the integument forms a large cavity, a pit, that receives the basal parts of the legs IV (" puits tØgumentaire" in Coineau 1964).The cuticle of these hollows is covered with dense villose integument similar to what has been observed in congeneric species (PT, Figs 1D View FIGURE , 2C) (cf. Coineau 1964; Feider, Vasiliu 1968).
Legs ( Figs 1A View FIGURE , 2A, 2D) — The legs are robust and sclerotized, the polygonal pattern ornamentation of the body is continued on the articles and discernible on femora and genua on the first pair of legs. The articles of the first pair of legs are thick, and not elongated as in the other genera of labidostommatids. However, asymmetry between right and left legs of the first pair attests that either the fossilisation process or optical properties of amber induced some deformation, the legs of the right side keeping more "natural" proportions. The third leg is missing on the left side. The alveolate ornamentation of the legs is similar to that of the shields, but locally it is resolved in cristules as on the edges of the genu and tibia of leg I ( Fig. 2A). The genu, is shorter than the telofemur and the separation of the femora in three articles is functionally reduced to basi- and [mesotelo] femora, due to "ankylosis" of the distal article of the femora ( Figs 2A, 2B) already described in the extant species ( Coineau 1964).
Tarsi as for the genus, tarsi II, III, IV are tridactylous, and tarsi of legs I bidactylous. The tarsi I are provided with long dorsal setae, and on the left leg, one of the two usual recumbent solenidia (ω) is clearly visible, and a spine which, by both the position and its shape, could be identified as the famulus ( Fig.5B, F View FIGURE ?).
Differential diagnosis — By the characters described here above, there is no doubt that this specimen belongs to the genus Eunicolina . The fossil species is distinct from the extant members of the genus by the supposed absence of frontal eye, reduced lateral eyes and by the lower number of pustules than in modern species. It clearly differs from E. nova ( Sellnick, 1931b), which has 20 to 35 pustules, by absence of neotrichy; from E. travei – by the absence of modified integument near the posterior trichobothria. On the ventral surface of the infracapitulum it clearly differs from both E. travei and E. nova, these latter species having respectively (ma) and (mb) and 4-5 or 7 pairs of infracapitular setae, while E. glaesi shows two pairs of infracapitular setae: (mb) and one other pair with discernible insertion marks.
Remark — The pustules of Eunicolina mites are distributed "on each side from the level of the leg II and extending uninterrupted caudad and absent from a short space of the posterior end of the dorsal shield" ( Greenberg 1952), but each species differs by their number ( Table 1). Berlese (1911) has noted the arrangement of the pustules in two "series" " tuberculi seriei marginalis 6 numero; marginalibus serie usque ad marginalis posticum producta, ad 25 numero ", i.e a total of 31 pustules, in E. tuberculata . E. travei presents the largest neotaxy, with more than 40 pustules on each side, in E. nova the number of pustules varies from 20 to 35 on each side ( Sellnick 1931b; Feider and Vasiliu 1968). The fourth species, the American E. porifera Greenberg, 1952 , from Virginia, averages 30 pustules on each side ( Greenberg 1952).
Etymology — from " glaesum ", term, used by Pliny the Elder (77) for amber ".../... certum est gigni in insulis septentrionalis oceani et ab germanis appellari glaesum, itaque et ab nostris ob id unam insularum glaesariam appellatam, .../... nascitur autem defluente medulla pinei generis arboribus .../...". Translation: It is well established that the amber is a product of islands in the Northern Ocean, that the Germans named "glaesum", and that, as a result, one of these islands was nicknamed "Glaesaria" by our (legion), .../... (Amber is) formed of a liquid seeping from the interior of a species of pine .../... [adapted from Woolley’s translation (1969)].
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.