
Microdipoena, BANKS 1895
|
publication ID |
https://doi.org/ 10.1111/zoj.12199 |
|
DOI |
https://doi.org/10.5281/zenodo.7007203 |
|
persistent identifier |
https://treatment.plazi.org/id/03832D77-1146-9261-FEE2-E43DFE1A5F36 |
|
treatment provided by |
Felipe |
|
scientific name |
Microdipoena |
| status |
|
MICRODIPOENA BANKS 1895 View in CoL View at ENA
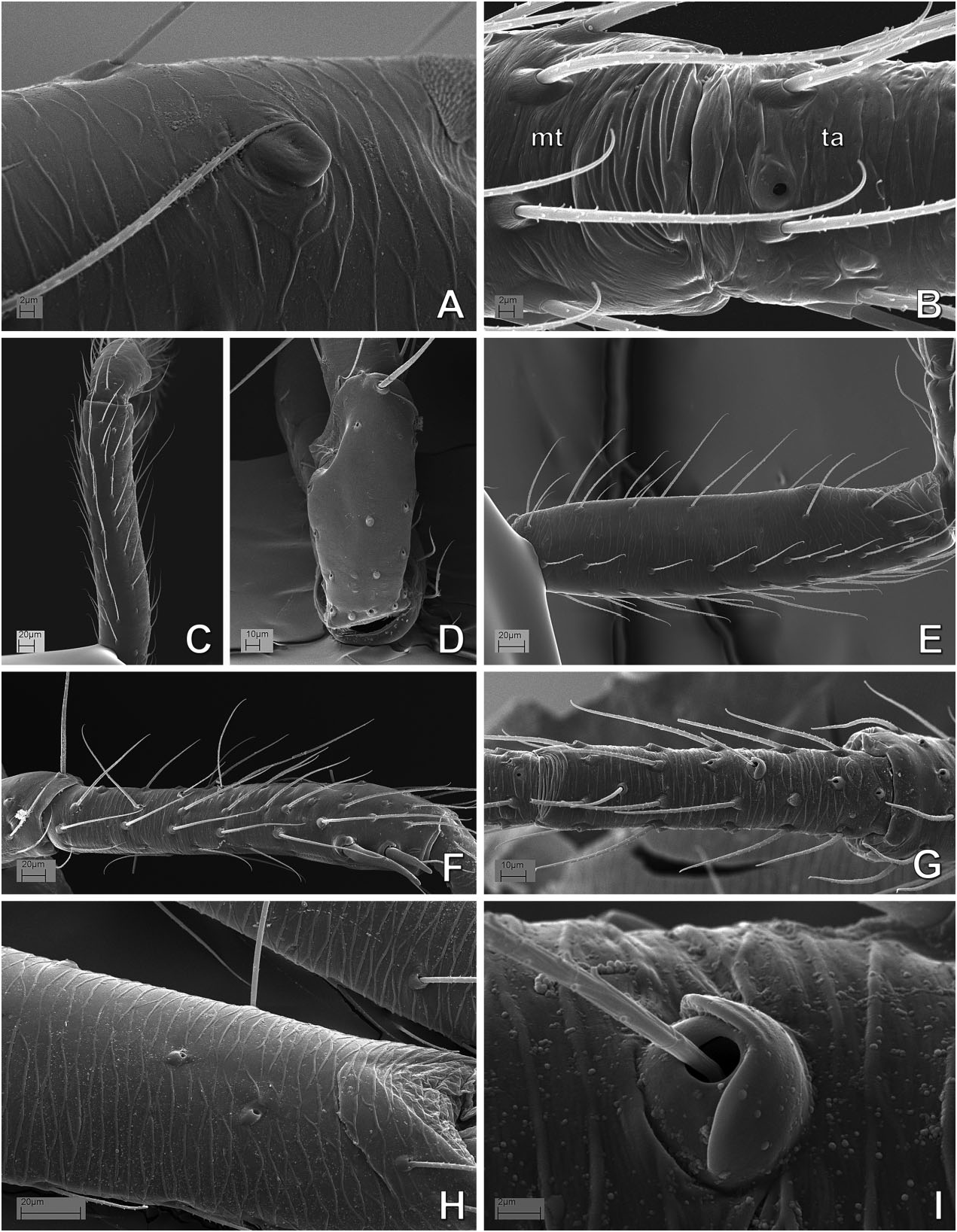
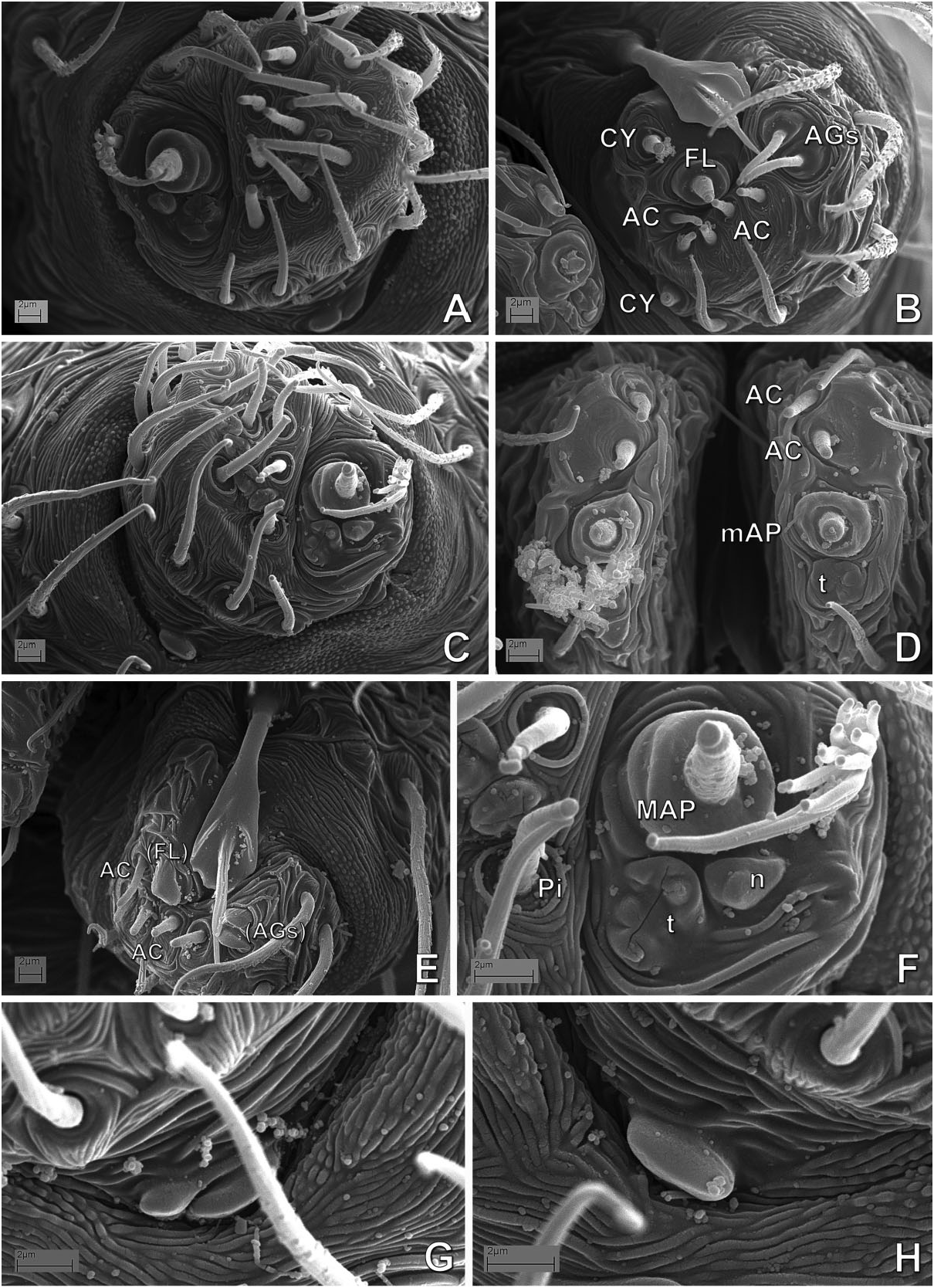
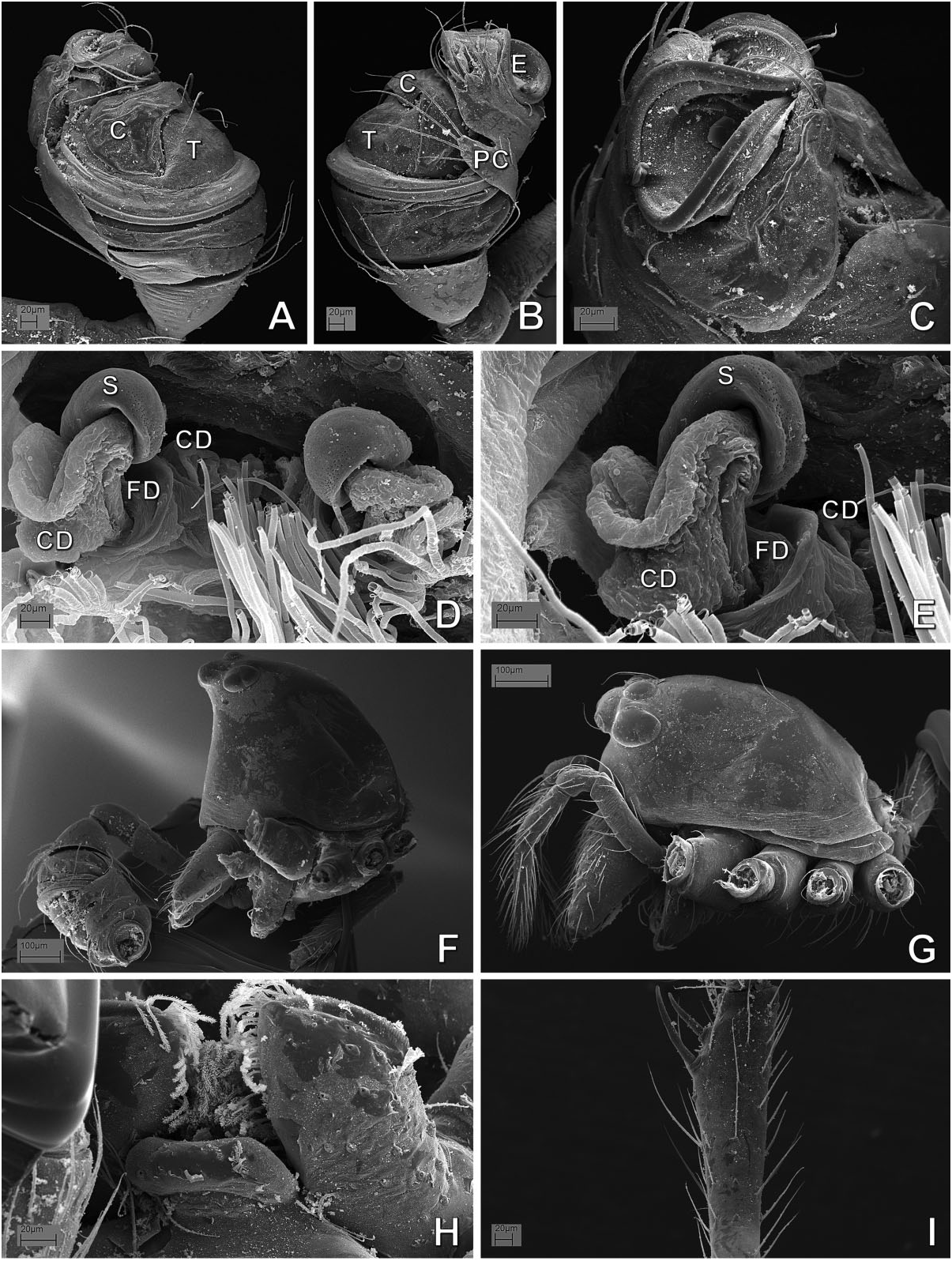
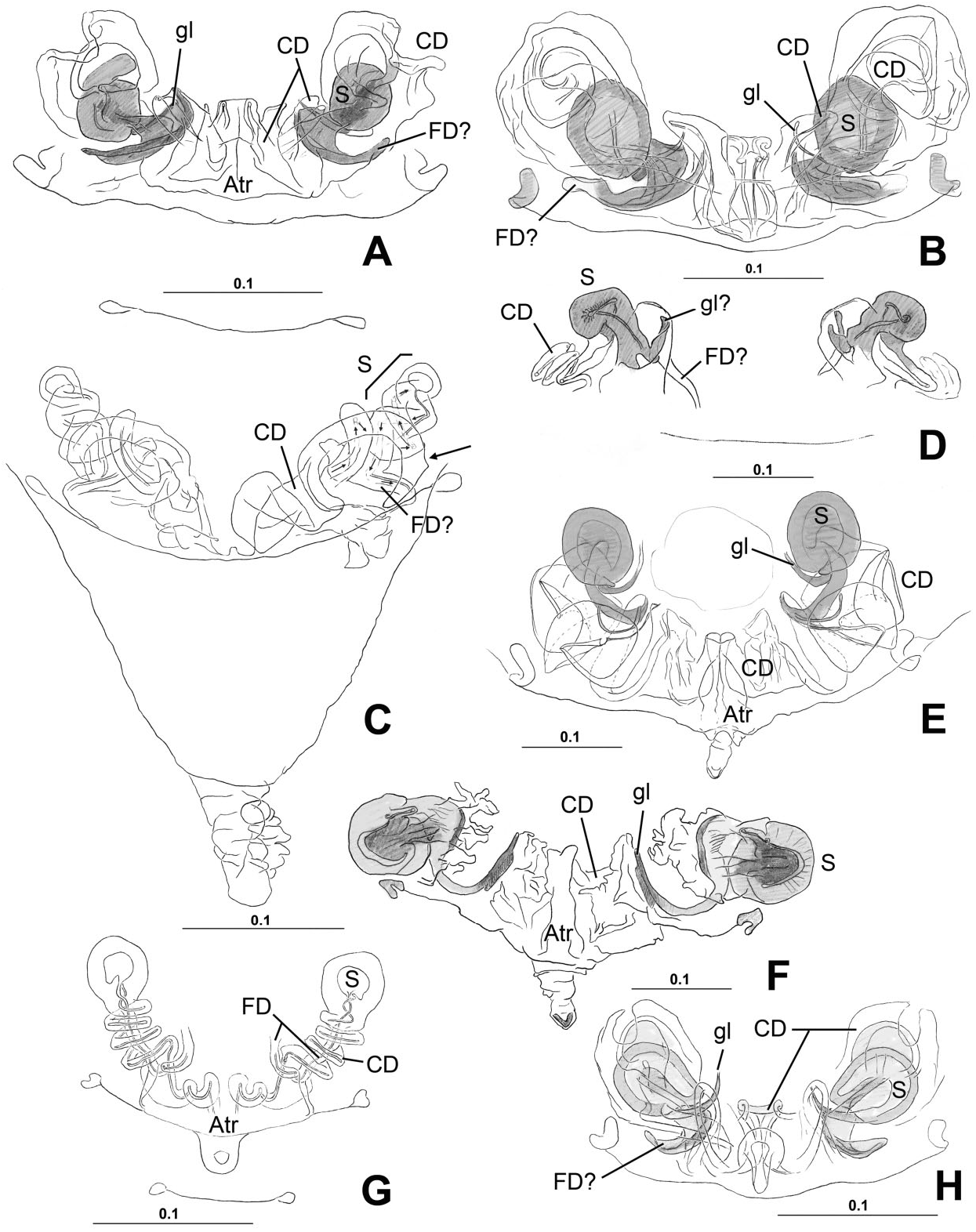
( FIGS 17–27 View Figure 17 View Figure 18 View Figure 19 View Figure 20 View Figure 21 View Figure 22 View Figure 23 View Figure 24 View Figure 25 View Figure 26 View Figure 27 , 129A, B, D–F View Figure 129 , 132 View Figure 132 , 141J–O View Figure 141 , 142A,B View Figure 142 : CLADE C125)
Mysmena Simon, 1895b: 149 View in CoL . Bishop & Crosby, 1926: 177. Levi, 1956: 8. Forster, 1959: 306. Kraus, 1967: 392. Gruia, 1977: 162. Shinkai, 1977: 326. Roberts, 1978: 932. Wunderlich, 1980b: 267; 1986: 222. Kasal, 1982: 75. Heimer & Nentwig, 1991: 306.
Microdipoena Banks, 1895: 85 View in CoL . Saaristo, 1978: 124–125 (rejected synonymy to Mysmena View in CoL by Bishop & Crosby, 1926: 177). Brignoli, 1980: 731 (rejected synonymy to Mysmena View in CoL by Bishop & Crosby, 1926: 177). Baert, 1984b: 608; 1985: 51; 1989: 29.
Anjouanella Baert, 1986: 265 (type species by monotypy A. comorensis Baert, 1986 , type material in MRAC, examined). New synonymy.
Mysmenella Brignoli, 1980: 731 (transfer from Mysmena View in CoL , type Mysmena illectrix Simon, 1895b , type material in MNHN, examined). Baert, 1984a: 240 (transfer from Mysmena View in CoL ); 1989: 32. Namkung & Lee, 1987: 46. Coddington, 1990: 19. Thaler & Noflatscher, 1990: 174. Namkung, 2002: 146; 2003: 148. Wunderlich, 2004: 1073 (considered a junior synonym of Mysmena Simon, 1894 View in CoL ). Yin et al., 2004: 80. Lee et al., 2004: 100. Trotta, 2005: 170. Ono, 2007: 170. New synonymy.
Type species
Microdipoena guttata Banks, 1895 View in CoL by original designation, type material in MCZ, examined.
Familial placement, composition, and re-circumscription
Our working phylogenetic hypothesis places Microdipoena sister to Brasilionata within the mysmenine clade C128, which also comprises Mysmeniola and MYSM-019-MAD. Microdipoena comprises four described species (Platnick, 2014) and under the current re-circumscription, 11 other described species are transferred here (a total of 15 described species). Microdipoena is here represented by seven described plus two undescribed species ( Fig. 161B View Figure 161 ; the latter two species are scored only for molecular characters): M. guttata , M. elsae , M. nyungwe , M. samoensis comb. nov. (from Mysmenella ), M. jobi comb. nov. (from Mysmenella ), M. illectrix comb. nov., M. comorensis comb. nov., Microdipoena -AToL-DR, and MYSM-030- MAD.
Monophyly, diagnosis, and synonymy justification The following combination of morphological synapomorphies is unique and therefore diagnostic for Microdipoena (and are shared among all Microdipoena representatives, unless noted): abdomen with a whitish ventral ring around the spinnerets ( Fig. 142A View Figure 142 ; except Anjouanella , with all ventral abdominal area lighter, Fig. 141J–L View Figure 141 ); males with two prolateral apical clasping spines on tibia I ( Figs 26C View Figure 26 , 27I View Figure 27 , 141K, L, O View Figure 141 ), thick embolus with an apical switch in the coiling direction ( Figs 18C, D, F View Figure 18 , 27C View Figure 27 , 132B, D, E View Figure 132 ; also in Brasilionata ), and with either a distal apophysis ( Fig. 18F View Figure 18 ) or a distal irregular membrane ( Fig. 27A– C View Figure 27 ; except in Anjouanella , without modifications), spermatic duct switchback SB I parallel, with the portions of the spermatic duct before and after the switch SB I run close with each other and with one pair of extra switches (SB III and IV, Fig. 132B–E View Figure 132 ); and small paracymbium ( Figs 17C View Figure 17 , 22G View Figure 22 , 27B View Figure 27 ). As most Microdipoena representatives in this data set were scored only for morphology, no molecular synapomorphies optimize at the node of this genus; however, its distal clade (clade C172), which includes the only sequenced species of this genus, is supported by 92 molecular synapomorphies. Previous diagnoses for Microdipoena s.s., Mysmenella , and Anjouanella are in agreement with the current diagnosis of the enlarged Microdipoena (see e.g. Banks, 1895; Brignoli, 1980; Baert, 1986).












No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Microdipoena
| Lopardo, Lara & Hormiga, Gustavo 2015 |
Mysmena
| Bishop & Crosby, 1926: 177 |
| Levi, 1956: 8 |
| Forster, 1959: 306 |
| Kraus, 1967: 392 |
| Gruia, 1977: 162 |
| Shinkai, 1977: 326 |
| Roberts, 1978: 932 |
| Wunderlich, 1980b: 267; 1986: 222 |
| Kasal, 1982: 75 |
| Heimer & Nentwig, 1991: 306 |
Microdipoena
| Saaristo, 1978: 124–125 |
| Bishop & Crosby, 1926: 177) |
| Brignoli, 1980: 731 |
| Baert, 1984b: 608 |
| 1985: 51 |
| 1989: 29 |
Mysmenella
| Baert, 1984a: 240 |
| Namkung & Lee, 1987: 46 |
| Coddington, 1990: 19 |
| Thaler & Noflatscher, 1990: 174 |
| Namkung, 2002: 146 |
| 2003: 148 |
| Wunderlich, 2004: 1073 |
| Yin et al., 2004: 80 |
| Lee et al., 2004: 100 |
| Trotta, 2005: 170 |
| Ono, 2007: 170 |