
Tartarocyon cazanavei, Solé & Lesport & Heitz & Mennecart, 2022
|
publication ID |
https://doi.org/10.7717/peerj.13457 |
|
publication LSID |
lsid:zoobank.org:pub:9FE7C271-9402-4062-B9B5-2087C8ACDC04 |
|
DOI |
https://doi.org/10.5281/zenodo.6791681 |
|
persistent identifier |
https://treatment.plazi.org/id/C7BE021C-6434-4715-AB89-63E9A64E6178 |
|
taxon LSID |
lsid:zoobank.org:act:C7BE021C-6434-4715-AB89-63E9A64E6178 |
|
treatment provided by |
Felipe |
|
scientific name |
Tartarocyon cazanavei |
| status |
sp. nov. |
Species Tartarocyon cazanavei nov. gen. & sp.
Figure 3 View Figure 3
ZooBank LSID. urn:lsid:zoobank.org:act:
Etymology. Dedicated to Mr Alain Cazanave, owner of the locality, who helped with the excavation during many years.
Diagnosis. Large size Amphicyoninae possessing a complete dental formula. The taxon is characterized by the following features: long diastemata between the premolars, low p2 and p3, absent anterior accessory cuspid on p4, large and individualized distal accessory cuspid on p4, and unreduced m2 and m3. The taxon differs from all the European amphicyonids from the Miocene by the individualization of the distal accessory cuspid from the main cuspid on p4 and the extreme reduction of the distal shelf and cingulid.
Specimen. MHNBx 2020.20.1 , right mandible bearing p2-p4, alveoli of i1-i3, c, p1, m1-m3 .
Measurements. Tables 1 View Table 1 & 2 View Table 2 .
Description. The mandible is mesiodistally elongated. Large diastemata are present between the canine, p1, p2, p3, and p4; the longest diastema is between the p2 and p3.
The symphysis is oval and nearly horizontally oriented; it is high and extends posteriorly up to the distal root of p2. A mental foramen lies beneath the p1-p2 diastema; it is in a high position on the mandibular ramus. The ramus of the mandible is shallower anteriorly than posteriorly, the highest portion being below the m3. The ventral margin of the ramus below the toothrow is relatively straight, but beneath the anterior extremity of the large, deep masseteric fossa it becomes convex. An incisura vasorum is present on the ventral margin of the mandible anterior to the angular process. The angular process is robust but very short; it projects medially. The mandibular condyle is at the level of the tooth row. It is cylindrical and mediolaterally elongate. The coronoid process is tall and distinctly oriented backwards; it arises at a 50 ◦ angle relative to the horizontal ramus. The posterior margin of the coronoid is vertical and straight, while the cranial margin is rounded. The masseteric fossa, on its labial side, is deep and wide. The mandibular foramen is relatively circular, standing at the level of the incisura vasorum, at mid-height between the base of the mandible and the level formed by the toothrow. The mandibular foramen opens midway between the m3 and the mandibular condyle.
The lower incisors are not preserved, but the alveoli of the i1, i2, and i3 are visible. Considering the size of the tooth sockets, the i3 seems to have been the largest and the i1 the smallest. The canine is also not preserved. It was ovoid in section and of large size. Its root extends in the mandible to between p2 and p3. The p1 is not preserved; a single alveolus is visible, but it appears that two, mainly fused, roots were present. The other teeth are two-rooted, except the m3, which is single-rooted. The p2 and p3 are very low in height. There is a prominent ridge on the mesial and distal margins of the main cuspid of these teeth. The main cuspid is low and located mesially, which results in an asymmetric morphology in lateral view. Mesial to the main cuspid, the lingual cingulid is thicker, but no individualized anterior cuspid is present. On p3 and p4, the distal shelf forms the widest part of the crown; it is less clear on p2. There is a short distal cingulid, but no cuspid is present. The p4 is distinctly longer and mediolaterally wider than the p2 and p3. However, the main cuspid remains low. The tooth is less asymmetric, the apex of the main cuspid being more mesiodistally centered. No real anterior accessory cuspid is present mesial to the main cuspid. A distal accessory cuspid is present: it is mostly individualized from the main cuspid. The distal accessory cuspid is mediolaterally centered. The distal cingulid is thin on the labial and lingual parts and is almost completely absent at the distal part; it does not form a distal shelf. The molars are not present, but the m1 was the largest tooth of the tooth-row. The m2 is larger than the m3.
Comparison. The premolars of the typical haplocyonines ( Haplocyon , Haplocyonoides , Haplocyonopsis ; de Bonis, 1966; Peigné & Heizmann, 2003; Morlo et al., 2020) differ from those of MHNBx 2020.20. 1 in being high ( i.e., high main cuspid) and in the loss of the p4 distal accessory cuspid. Like the typical haplocyonines, the premolars of Gobicyon serbiae (MN6) differ from those of MHNBx 2020.20. 1 in being high. Moreover, the p2 and p3 of G. serbiae possesses an individualized and large distal accessory cuspid. Additionally, typical haplocyonines and Gobicyon have a short toothrow lacking diastemata. These amphicyonids are thus relatively short-snouted compared to the taxon from Sallespisse.
All the thaumastocyonines differ from MHNBx 2020.20. 1 in having relatively shorter diastemata between the premolars. The p2 and p3 preserved on MHNBx 2020.20.1 are similar to those of the oldest thaumastocyonines ( Ysengrinia , Crassidia ) in being low ( i.e., their main cuspid is noticeably lower than the p4 main cuspid). The p4 of MHNBx 2020.20.1 also shares with the thaumastocyonines the presence of a strong distal accessory cuspid ( Fig. 4 View Figure 4 ); the youngest thaumastocyonines ( e.g., Agnotherium, Ammitocyon ) shares with the p4 of MHNBx 2020.20.1 the reduced distal shelf and cingulid ( Fig. 4 View Figure 4 ). However, the p4 of the thaumastocyonines differs from that of MHNBx 2020.20. 1 in having a leaning backward p4 main cuspid ( Fig. 4 View Figure 4 ). The youngest thaumastocyonines Ammitocyon and Agnotherium moreover, differ from MHNBx 2020.20. 1 in having no p1, p2, and p3 ( Morlo et al., 2020; Morales et al., 2021a). Compared to the fossil from Sallespisse, the thaumastocyonines have a reduced m3 relative to m1; the youngest thaumastocyonines ( Thaumastocyon .
Ammitocyon, Agnotherium ) have even reduced m2 relative to m 1 in addition to lacking m3 ( Morlo et al., 2020; Morales et al., 2021a). As a consequence, MHNBx 2020.20.1 differs in having more developed premolars, a mesially elongated snout ( i.e., diastemata between the premolars), and less reduced postcarnassial molars.
Three amphicyonines are regarded to be separate from those recorded in the Miocene: Ictiocyon, Dehmicyon , and Pseudarctos ( Ginsburg, 1992; Morales et al., 2021b). They are all included among Pseudarctini ( Morales et al., 2021b). These small amphicyonids are short-snouted ( i.e., short diastemata between the premolars) and the p2 and p3 are distinctly taller than on MHNBx 2020.20.1. Moreover, the distal accessory cuspid on p4 is reduced to lost in Dehmicyon, Ictiocyon , and Pseudarctos ( Ginsburg, 1992; Morales et al., 2021b) ( Fig. 4 View Figure 4 ).
The hypercarnivorous Magericyon ( Peigné et al., 2008) , which belongs to the tribe Magericyonini ( Morales et al., 2021b) differs from MHNBx 2020.20. 1 in the absence of p2, in having a single-rooted p3, a p4 relatively shorter compared to the m1 ( Table 3 View Table 3 ) and in the absence of a distal cuspid on p4 ( Fig. 4 View Figure 4 ). The genus Pseudocyon is probably close to Magericyon according to Morales et al. (2021b). MHNBx 2020.20.1 is similar to the species of Pseudocyon in the presence of very long diastemata between the premolars and of low p2, p3. However, the p4 is relatively mesiodistally shorter (compared to the m1) in the Pseudocyon species than in MHNBx 2020.20.1; moreover, the distal part of the p4 of Pseudocyon is widened compared to that of the p4 of MHNBx 2020.20.1 ( Fig. 4 View Figure 4 ).
The Miocene Amphicyonini Cynelos , Amphicyon , Megamphicyon , Euroamphicyon, Paludocyon, and Heizmannocyon share with MHNBx 2020.20.1 the presence of very long diastemata between the premolars, the presence of low p2, p3, and p4, and the unreduced m3 (the m3 indeed tends to reduce and is even absent in hypercarnivorous amphicyonids; Table 3 View Table 3 ) ( Kuss, 1965; Peigné & Heizmann, 2003; Viranta, 1996). Despite sharing a characteristically slender ramus of the mandible, the p4 of MHNBx 2020.20.1 differs from that of the Cynelos species by the absence of an anterior accessory cuspid (even if this structure is not individualized in Cynelos ) and a much more reduced distal shelf ( Fig. 4 View Figure 4 ). The case of Cynelos is interesting because its p4 does not display a widening of its distal part; in this regard, its p4 is similar to that of MHNBx 2020.20. 1 in occlusal view ( Fig. 4 View Figure 4 ). MHNBx 2020.20.1 shares with the species of Paludocyon, Amphicyon, Heizmannocyon , Megamphicyon , and Pseudocyon the reduction of the anterior accessory cuspid compared to Cynelos . However, the distal shelf of the p4 is more developed in these amphicyonines than in MHNBx 2020.20.1 and none of the above-mentioned species has a p4 that displays a distal accessory cuspid separated from the main cuspid as it is on the p4 of MHNBx 2020.20.1. Moreover, these amphicyonine genera (see Megamphicyon carnutense and Paludocyon bohemicus in Morales et al., 2021b) possess a p4 that is characterized by a widening of the distal part. Additionally, the mandible of Amphicyon and Megamphicyon appears more massive than that of MHNBx 2020.20.1 ( Kuss, 1965; Peigné & Heizmann, 2003; Viranta, 1996; Fig. 4 View Figure 4 ).
Table 3 ( continued)
A canine has been described from the locality of Rimbez ( France, MN5), a locality that is located 100 km to the north-west of Sallespisse ( Ginsburg, 1967); this locality is the closest one that has provided a Miocene amphicyonid specimen .
This canine has been referred to Pseudocyon sansaniensis , an Amphicyonidae of similar size to MHNBx 2020.20.1. It is at the moment impossible to compare this canine with MHNBx 2020.20.1, but one can note that this tooth is close in size to the alveolus of the canine of MHNBx 2020.20.1. Despite that the canine is much older than MHNBx 2020.20.1, one can imagine that the taxon from Rimbez could also be closely related to the taxon from Sallespisse.
To conclude, the fossil from Sallespisse shows striking similarities with the much older and clearly smaller Cynelos , a genus not yet known from the Middle Miocene of Europe ( i.e., presence of long diastemata between the premolars, unreduced premolars and m3, low p2 and p3, no widening of the distal part of the p4). The general morphology of the p4 remains relatively stable within the Amphicyoninae having a distal accessory cuspid more or less individualized and a distal shelf present ( Fig. 4 View Figure 4 ). MHNBx 2020.20.1 presents an unique morphology among the Amphicyoninae in having an individualized distal accessory cuspid on p4 and a distal shelf extremely reduced, extending the morphological range of the p 4 in this subfamily ( Fig. 4 View Figure 4 ). Therefore, we erect the new genus and species Tartarocyon cazanavei nov. gen. & sp. for MHNBx 2020.20.1.
One can think that the basis for erecting a new taxon on this fragmentary material is not strong. However, because we are unable to discern which genus it most closely resembles and that given the originality of the morphology of the p4, we hypothesize that this specimen represents a new genus that will be further confirmed or not by future finds.
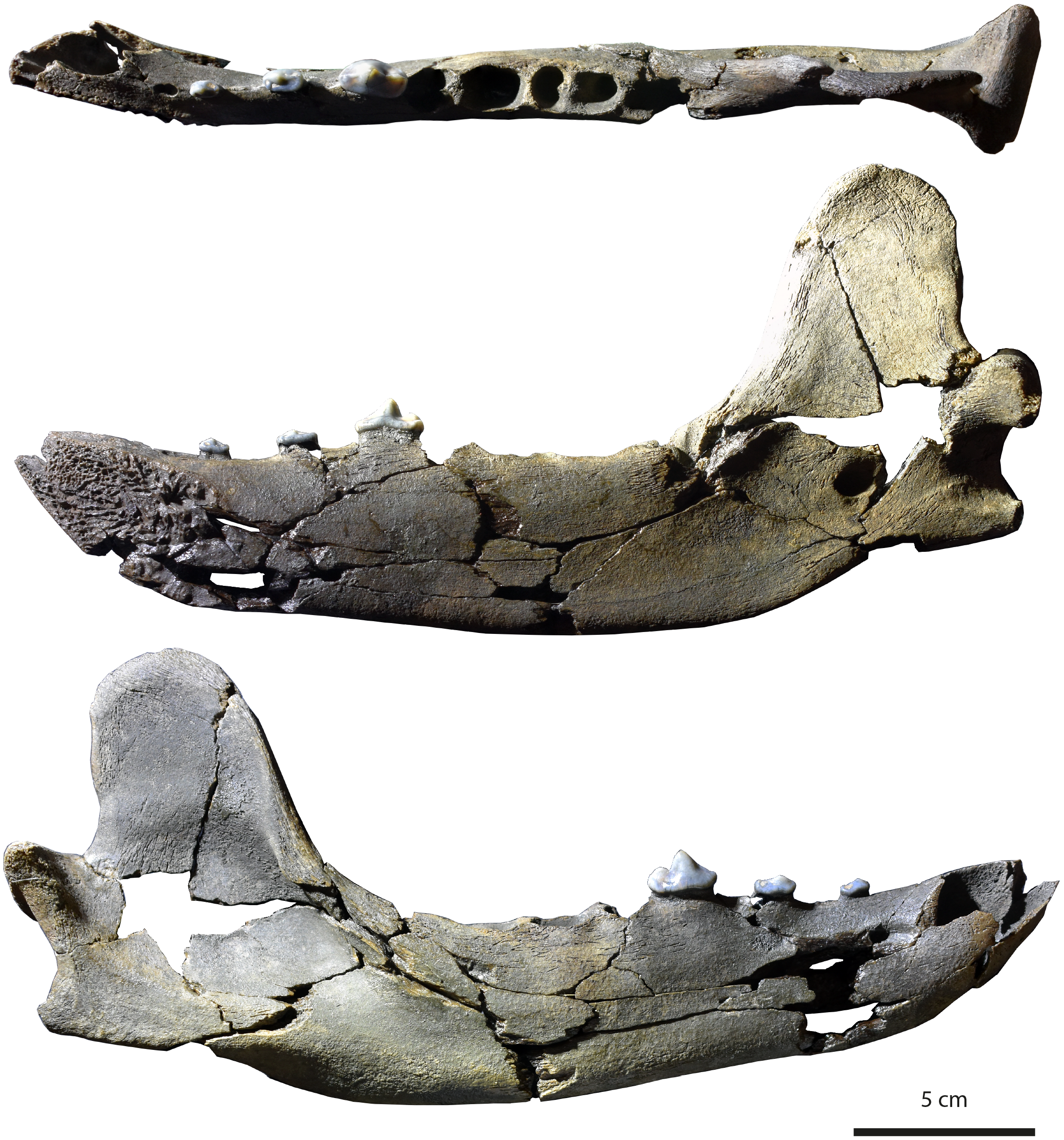
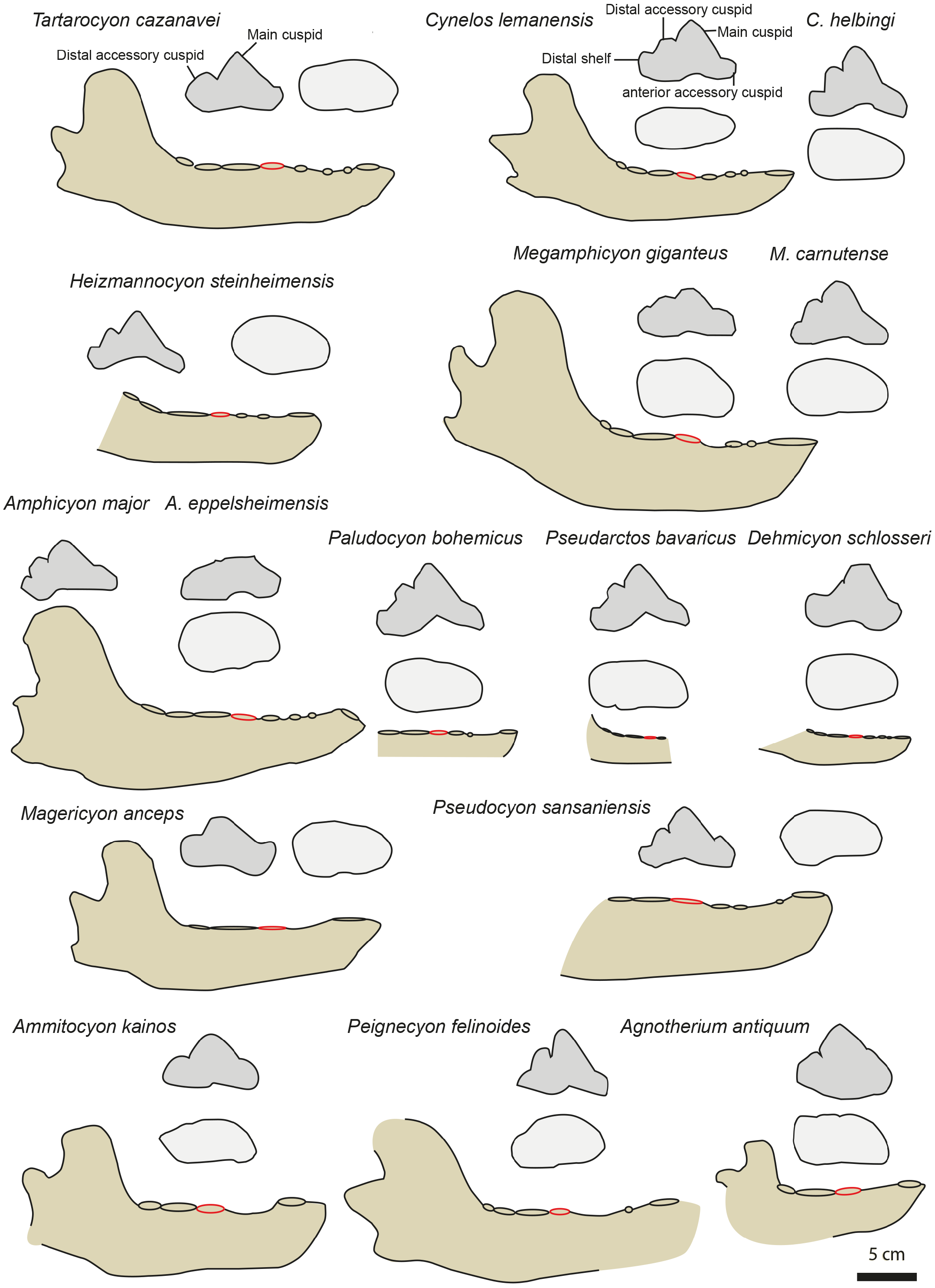
Table 1 Measurements of the teeth of the holotype (MHNBx 2020.20.1) of Tartarocyon cazanavei nov. gen. & sp. from Sallespisse (MN7/8).
| Tooth locus | Length | |
|---|---|---|
| i1 | 7.58* | 3.19* |
| i2 | 9.88* | 5.02* |
| i3 | 11.51* | 5.15* |
| c | 18.02* | |
| p1 | 7.87* | 3.86* |
| p2 | 8.27 | 4.63 |
| p3 | 11.14 | 6.35 |
| p4 | 18.58 | 9.67 |
| m1 | 34.30* | 13.88* |
| m2 | 24.26* | 14.22* |
| m3 | 17.21* | 11.93* |
Table 2 Several measurements of the teeth and mandible of the holotype (MHNBx 2020.20.1) of Tartarocyon cazanavei nov. gen. & sp. from Sallespisse (MN7/8).
| Length p1-p4 | 69.94 |
|---|---|
| Length m1-m3 | 78.67 |
| MD below p2 | 39.69 |
| MD below m1 | 48.97 |
| MD below m3 | 53.25 |
Notes.
MD, Mandible height.
Table 3 Ratios estimated based on premolars and molars for several amphicyonines and thaumastocyonines known in the Miocene of Europe.
| Taxon | Stratigraphic | Ratio | Ratio | Ratio | Ratio | Ratiov |
|---|---|---|---|---|---|---|
| distribution | Lp2/Lm1 | Lp3/Lm1 | Lp4/Lm1 | Lm2/Lm1 | Lm3/Lm1 | |
| Cynelos lemanensis MNHNL-La85 | MN1-MN2 | 0.43 | 0.67 | 0.63 | ||
| Crassidia intermedia SMNS 46684 | MN1-MN2 | 0.47 | 0.43 | 0.63 | 0.58 | 0.38 |
| Ysengrinia gerandiana FSL 213828 | MN1-MN2 | 0.44 | 0.47 | 0.62 | ||
| Cynelos rugosidens BSP-1881-IX-14, 581 | MN2 | 0.67* | 0.65 | 0.42 | ||
| Peignecyon felinoides TU 7391147 | MN3 | 0.55 | 0.49 | |||
| Megamphicyon carnutense Fs 6953 | MN3 | 0.35 | 0.53 | 0.59 | 0.71 | |
| Cynelos helbingi BSP-II-1937-12293 | MN3-MN4 | 0.57* | 0.64 | 0.39 | ||
| Ictiocyon socialis Ginsburg ( 1992, p. 311) | MN3-MN4 | 0.41 | 0.51 | 0.62 | 0.72 | 0.42 |
| Ysengrinia depereti MSNO. 785 | MN3-MN4 | 0.25 | 0.43 | 0.48 | 0.62 | 0.34 |
| Dehmicyon schlosseri BSP 13562 | MN3-MN5 | 0.37 | 0.48 | 0.61 | 0.59 | 0.37 |
| Paludocyon bohemicus NM-PV 11723 | MN3-MN5 | 0.43 | 0.49 | 0.59 | 0.65 | 0.37 |
| Pseudocyon sansaniensis MNHN.F.Sa 207 | MN3-MN9 | 0.28 | 0.29 | 0.51 | 0.6 | |
| Tomocyon grivense UCBL-FSL 213797 | MN3-MN9 | 0.6 | ||||
| Megamphicyon giganteus Specimen from Vienna & Basel SO6521 ( Hunt Jr, 2003, table 4.7) | MN4-MN7/8 | 0.3 | 0.42 | 0.58 | 0.71 |
( continued on next page)
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SubOrder |
Caniformia |
|
Family |
|
|
Tribe |
Amphicyonini |
|
Genus |