
Pachynolophus ruscassierensis, Remy & Krasovec & Lopez & Marandat & Lihoreau, 2019
|
publication ID |
https://doi.org/10.5252/geodiversitas2019v41a13 |
|
publication LSID |
urn:lsid:zoobank.org:pub:56DC3958-1615-45E9-91FF-8C47158A82FD |
|
DOI |
https://doi.org/10.5281/zenodo.3705062 |
|
persistent identifier |
https://treatment.plazi.org/id/F10B45D3-0CC9-4CD4-8E38-0C568BBC7038 |
|
taxon LSID |
lsid:zoobank.org:act:F10B45D3-0CC9-4CD4-8E38-0C568BBC7038 |
|
treatment provided by |
Valdenar |
|
scientific name |
Pachynolophus ruscassierensis |
| status |
sp. nov. |
Pachynolophus ruscassierensis n. sp.
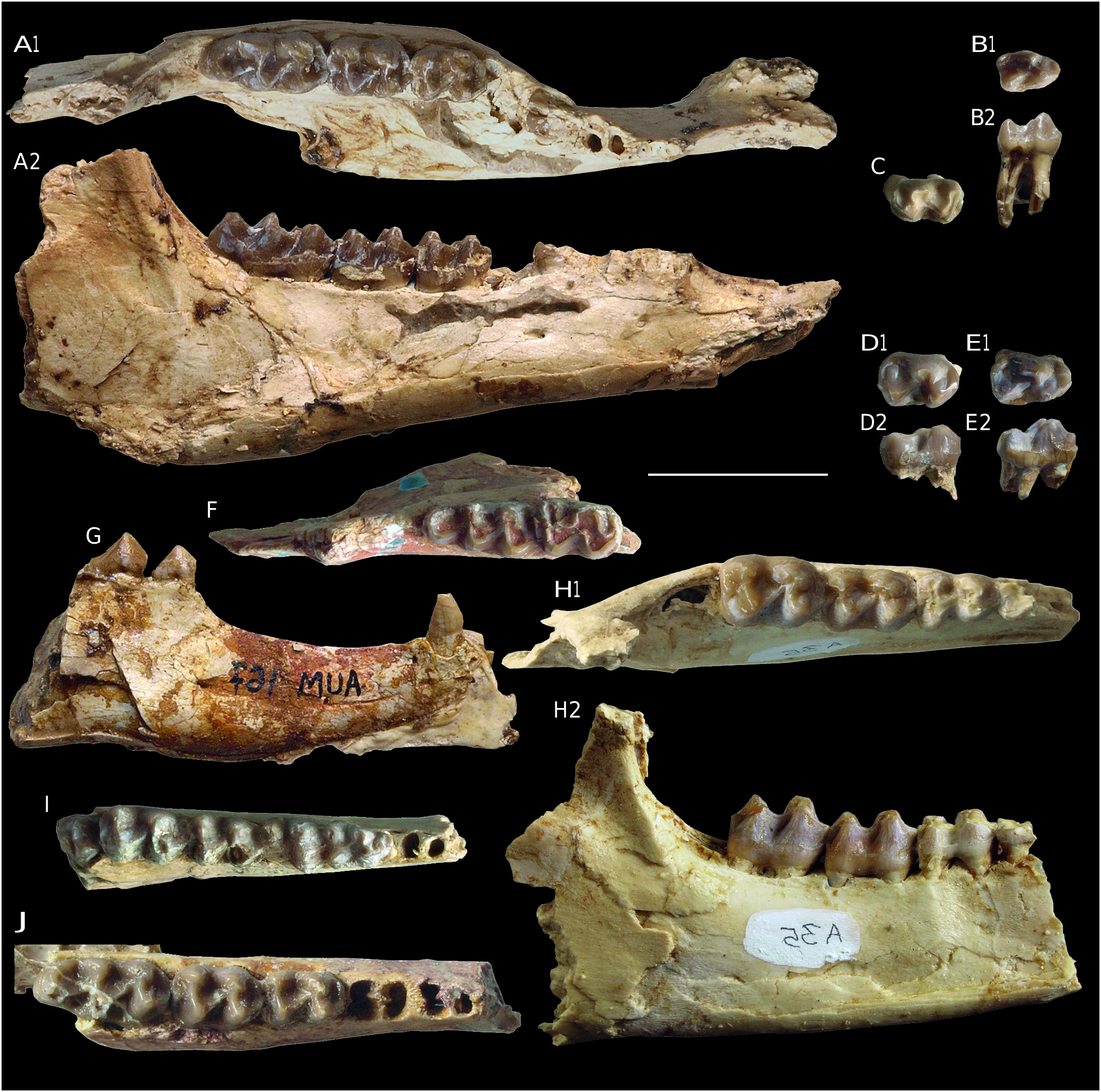
( Figs 8 View FIG , 9 View FIG )
TYPE SPECIMEN ( HOLOTYPE). — UM-AUM 231 fragmentary skull preserved from nasal opening up to zygomatic arch, with right (alv. C/-P1/)-P2/-M3/ and left (alv. C/, P2/)-P3/-M3/ ( Fig. 8B View FIG ).
TYPE LOCALITY. — Aumelas, Hérault department, France.
ETYMOLOGY. — From the geographic locality; in Occitan language, “ruscassier” evokes a place covered with trees having rough bark.
MATERIAL. — AUM 48, left M1/; AUM 70, left P3/; AUM 138, right maxillary fragment with P4/-M2/; AUM 176, right M1/; AUM 186, left DP3/; AUM 187, left M1/; AUM 189, left M2/; AUM 191, right M1/; AUM 194, left M1/; AUM 197, M1/; AUM 210, right maxillary fragment with M3/; AUM 211, right M2/; AUM 216, left maxillary fragment with M1/-M2/; AUM 224, left M3/; AUM 235, left M1/ (cast); AUM 304, right M3/; AUM 321, left maxillary fragment with (M1/)-M2/; AUM 324, left maxillary fragment with M3/; AUM 1544, palate with right (alv. C/)-P1/, M1/-M3/ and left P1/, P4/-M3/; AUM 1545, AUM 1546, AUM 1564, left M1/-M2/; AUM 1552, left maxillary fragment with P2/- (M3/); AUM 1553, left maxillary fragment with P3/-M2/; AUM 1554, right maxillary fragment with P4/-M2/; AUM 1555, right M2/; AUM 1563, right maxillary fragment with M2/-M3/; AUM 1565, left M2/; AUM 1566, right P3/; AUM 1567, left P4/; AUM 1618, right maxillary with DP2/-DP4/-M1/; AUM 1619, fragment of dislocated skull with right M1/-M2/; AUM 1628, left P3/-(M2/) (compositeṙ); AUM 1632, right P3/; AUM 1634, left M3/. AUM 161, left mandible with M/1-M/3, alveoli of P/1-P/4; AUM 167, mandible with symphysis, left P/1- P/2 and both canines; AUM 169, right mandible fragment with M/1-M/3, alveoli of P/3-P/4; AUM 177, left mandible fragment with M/1; AUM 178, right P/3; AUM 185, right M/2; AUM 217, right M/2; AUM 305, left M/2; AUM 1549, right DP/4; AUM 1556, right mandible with (alv. DP/2)-DP/3-M/2; AUM 1558, right M/2; AUM 1559, left M/1; AUM 1562, AUM 1612, right P/2; AUM 1568, right mandible with M/2-M/3; AUM 1569, right M/3; AUM 1570, left M/2- (M/3); AUM 1571 right fragment with M/3; AUM 1575, right M/1; AUM 1578, AUM 1637, right P/3; AUM 1579, right P/1; AUM 1582, AUM 1621, AUM 1639, right M/1; AUM 1605, left mandible with ( DP /3)-DP/4-M/2; AUM 1623, left M/1; AUM 1629, AUM 1636, AUM 1640, AUM 1642, AUM 1643, right P/4; AUM 1630, left DP/4; AUM 1631, AUM 1635, left (M/3); AUM 1638, right M/3; AUM 1641, right M/1; AUM 1657, left M/2.
DIAGNOSIS. — Rather large Pachynolophus species, LRDJ = 51- 53 mm; estimated weight about 21 kg. Buno-lophodont dentition. Ectoloph of upper molars fairly flat, devoid of mesostyle and usually without pseudomesostyle. Deeply hollowed centrocrista. Relative surface of the premolar area not greatly reduced for a Pachynolophus , estimated at 44% of that of molars. Transversally elongated upper premolars, but with a lingual contour not specially narrowed. Rather straight lophids on lower molars; the transverse ones dug at midway. Labial crest of the M/3 hypoconulid connected to the mid hypolophid. Relatively thick and high cingula, almost wholly surrounding upper premolars, only missing on lingual side of lower molars. DPC of variable length but generally less elongated than in most other Pachynolophus .
DIFFERENTIAL DIAGNOSIS. — Larger than most Pachynolophus species, except Pa. cayluxi. More lophodont dentition than that of Pa. eulaliensis and Pa. cesserasicus , but much less than that of Pa. lavocati and Pa. zambranensis. Centrocrista of upper molars more dug than those of Pa. bretovensis and Pa. garimondi . Greater relative surface of the premolar area than that of Pa. molipontiensis, Pa. livinierensis , Pa. cesserasicus and Pa. lavocati . Less transversally elongated P4/ than in Pa. cayluxi, Pa. gaytei and Pa. cesserasicus with a less narrow lingual outline. Presence of a distinct metaconule on P4/, contrary to Pa. boixedatensis and M3/ not so large. Postero-lingual basin of P4/ less broad than that of Pa. bretovensis and Pa. garimondi . Thicker and higher cingula than in these latter species, wider also than in Pa. duvali .
DESCRIPTION
Upper cheek teeth
The type-specimen AUM 231 is in very good condition; only the M1/ bears notable signs of wear. It is a medium-sized Pachynolophinae (LRDJ = 51.3 mm), slightly smaller than Propalaeotherium sudrei but larger than most hyracotheres. Dimensions of the whole material reported to the taxon suggest a body weight of about 21 kg following the equations in Janis (1990) (Appendix 8), with coefficients of size variability (V) mostly <10% ( Table 3). The dentition is characterized by its heterodonty with an increasing size from P2/ to M3/. The crowns are rather low. The IH index of AUM 231 reaches only 0.37, but it is higher on other molars (AUM 324, AUM 191, AUM 194), up to 0.48-0.53 (Appendix 12).
The molar outlines become more trapezoidal backward from M1/ to M3/, narrowed distally, and only slightly transversally elongated. In spite of a global homogeneity in shape, some morphological variations are noticed. Ectoloph of M1/ can be less oblique than on the type-specimen (AUM 187), or on the contrary, more oblique (AUM 1564). M1/ can also be narrower distally with a labially shifted hypocone (AUM 176). Similarly, M3/ can have a more oblique ectoloph than on the holotype, with a recessed hypocone (AUM 224, AUM 324). The ectoloph is rather flat, though bearing a stout labial bulge on paracone. The metacone is slightly tilted inside and its labial bulge is less protruding. The centrocrista can be deeply notched (AUM 231, AUM 194) or only slightly dug (AUM 191, AUM 324, AUM 321). The parastyle is fairly strong and prominent on AUM 324, with the metastyle notably marked on M3/. The labial cingulum is not very salient although continuous and rather high on AUM 231. It is very thin and/or a bit interrupted on some specimens (AUM 138, AUM 187, AUM 191, AUM 1545). There is no mesostyle, although a slight crest is observed on mid ectoloph of the right M2/ of AUM 231.
The protoloph is directed towards parastyle and connects the preparacrista. The paraconule is elongated and linguodistally expanded. The groove separating paraconule and protocone is not deep on AUM 231, especially on M1/. It can be more dug (AUM 176, AUM 321) or conversely, hardly noticeable (AUM 210, AUM 189, AUM 191, AUM 1545). The metaloph, on which any individualized metaconule cannot be distinguished, is connected to the mesio-lingual side of the metacone. The internal outline of molars is hollowed at the level of the medivallum. The hypocone is only slightly receded labially with respect to the protocone. The whole inner part of molars are surrounded on AUM 231 by relatively well marked cingula, which weaken from M1/ to M3/ at lingual side and are interrupted on cusps. On other specimens, the lingual cingulum is thick (AUM 224, AUM 304), restricted to medivallum (AUM 324, AUM 187, AUM 321, AUM 1554) or even missing (AUM 210, AUM 191, AUM 138, AUM 235). The distal outline is slightly convex on most M3/, without hypostyle. On some specimens, it is more widened distally, without any raised cusp (AUM 324) or with a small typical hypostyle (AUM 224).
The surface of the premolar series relative to that of molars is low, as usual in Pachynolophus ( Remy 1972) . On AUM 231, the ratio SP/SM is estimated at 44 (Appendix 21) and the PMI at 65.7.
P4/ is transversely elongated; the labial bulge of paracone and metacone is very marked and the centrocrista of ectoloph less notched than on molars. P3/ has an identical shape, but it is slightly less transversally elongated, with closer outer cusps and less notched centrocrista. As molars, the premolars exhibit some morphological variability. Indeed, some premolars differ markedly from homologous teeth of the holotype. Thus, some P3/ and P4/ have a pronounced concavity of the labial outline ( AUM 1553 , AUM 1628). There is a strong labial cingulum on the P3/-P4/ of AUM 231, continuous and extended by a poorly developed parastyle. The latter can still be weaker ( AUM 1544 , AUM 1554 [P4/], AUM 1552, AUM 1566 [P3/]). The protocone of P3/-P4/ is generally axially located. It is slightly shifted forward on AUM 231, and even more forward on some P3/ ( AUM 1552 ). The paraconule of these teeth is bulbous on AUM 231, rather separated from the protocone and clearly lower. It is weaker with the groove between paraconule and protocone being shallower on other specimens ( AUM 1554 AUM 1567, AUM 1628). The rounded metaconule is labially extended by a thin rectilinear postprotocrista ridge oriented toward the centrocrista on AUM 231. The transverse lophs are more V-shaped with less distinct conules on AUM 1552 and AUM 1553. On the holotype, a thick and continuous cingulum surrounds the whole inner part of P3/-P4/ up to the ectoloph, but without any enlargement of a posterior basin unlike in some P3/ ( AUM 1552 , AUM 1553). The lingual cingulum is weaker and interrupted at protocone level on some P4/ ( AUM 1554 , AUM 1567) or P3/ ( AUM 1552 , AUM 1553). The P 3/ ( AUM 70 ), which displays a particularly weak and low metaconule, has a forward shifted protocone and lacks lingual cingulum .
A P2/ is preserved on the holotype. It is a small ovoid tooth, narrow mesially and wide distally. It has two very close labial cusps with the metacone hard to individualize and lower than the paracone. The other known P2/ (AUM 1552) is a bit smaller and more rounded in shape. Its high labial cusp is extended distally by a crest devoid of any hint of an incipient metacone. The ectocingulum is faint and the styles poorly defined on AUM 231. The oblique lingual cingulum is enhanced by a small elongated protocone. In the distal basin, a tiny and very low crest evokes a faint preprotocrista. On AUM 1552, the protocone is more individualized and there is no crest in the worn basin.
The P1/ are preserved on AUM 1544. They are simple narrow teeth with a prominent cusp barely shifted rostrally and a surrounding cingulum. It seems that there was place for a short diastema between P1/ and P2/. The presence of a P1/ is also attested on AUM 231 by two alveoli indicating a tooth as long but narrower than P2/. There was no P1/-P2/ diastema on this specimen. The alveolus of upper canine indicates a large and rather narrow tooth.
The deciduous dentition is known by an upper jaw (AUM 1618), whose teeth are unfortunately fractured and crumbled, and by a DP3/ (AUM 186). The DP4/ is molarized, has a high and rather protruding parastyle and lacks a mesostyle. The groove between paracone and protocone appears rather deep. The hypocone is less lingual in position than the protocone. The lingual cingulum is narrow and interrupted on hypocone. The DP3/ are also molariform but with a slightly oblique mesio-lingual outline. On AUM 186, the protocone is shifted labially relative to hypocone and slightly lower, with a deep lingual notch between protocone and hypocone. The paracone-protocone groove is shallower than on AUM 1618. The metaloph is curved on AUM 186 due to the presence of a small accessory distal cusp. The cingulum is thick and high, surrounding the tooth and only slightly broken at the paracone. DP2/ is an elongated narrow triangular tooth. The paracone is prominent with a close low metacone. The parastyle is welldeveloped like an anterior cusp. The inner part of the tooth is chipped. DP1/, which is not preserved, had two roots.
Lower cheek teeth ( Fig. 9 View FIG )
The lower molars are characterized by rather pointed crescents. The lophids are rather straight and lophodont. The transverse ones appear to be dug midway when unworn. The trigonid is rather short, even shorter than talonid. The preprotocristid is lowered, and curved mesially, without any trace of paraconid. The metaconid is clearly splitted. The metalophid is connected between metaconid and metastylid. The hypoconulid is well developed on the middle of distal edge of M/1 and M/2. The labial crest of hypoconulid of M/3 reaches the middle of hypolophid and the curved lingual crest bears small knobs (AUM 161, AUM 169). The ectocingulum of molars is usually well marked, relatively high (about 2 mm for a total unworn cusp height of 6 mm on AUM 161) and fairly continuous (AUM 161, AUM 217), but can be weaker and more or less interrupted on cuspids (AUM 169, AUM 185, AUM 1568, AUM 1605). Not any lower molar bears a lingual cingulum.
The lower premolars are known only by isolated teeth. Those that we interpret as P/4 (see material) are rectangular and generally not narrowed mesially. The trigonid is molariform with a split metaconid (AUM 1640). The mesial crest of the protoconid is lowered; on AUM 1640 it terminates by a small, split paraconid. The talonid is slightly lower than the trigonid. The hypoconid is shifted labially. Although there is no entoconid, the hypoconid is sometimes extended by a low lingual cristid, a weak clue of an incipient cuspid (AUM 1642). The ectocingulum is generally weak, sometimes restricted to the medivallum (AUM 1629, AUM 1643). P/3 has an identical morphology, but the tooth tapers anteriorly, thus the trigonid is narrower than the talonid and somewhat more flattened (AUM 178). Like on P/4, the hypoconid is shifted labially, without any entoconid. The ectocingulum is rather weak. Three P/2 could be assigned to the species. They are simple teeth with one prominent cuspid (protoconid) slightly shifted rostrally. A parasagittal mesial ridge and a slightly linguo-distal one represent the relicts of the trigonid crescent. On AUM 1612, both ridges end with a very small knob. There is no true talonid, only another low parasagittal crest behind protoconid, terminated by a very small tubercle equivalent to a vestigial hypoconid. The ectocingulum is noticeable, but stopped on protoconid. P/1 (AUM 167), is an even more simple, bi-rooted tooth. The canines, also present on that specimen, are small suggesting it belongs to a female. The most complete canine (on the right) is eroded by a large distal surface of wear.
DP/4 is molariform but can be slightly narrow mesially (AUM 1605). It differs from the molars by the lower crown height and the thinness of the enamel (AUM 1630). The paralophid is devoid of a paraconid and the metaconid is clearly split. The talonid is as high as the trigonid and the entoconid as large as the hypoconid. The presence of an hypoconulid is noticeable. The ectocingulum is weak and interrupted on cuspids. DP/3 is narrow mesially but the crescent of the trigonid is fully developed (AUM 1556). An accessory small cuspid is observed on the lingual side of the mesial cristid of the protoconid. The talonid is wide and the entoconid is as high as the hypoconid like on DP/4.
Other anatomical data
The length of the post-canine diastema (DPC) appears variable in the sample. On a maxilla (AUM 231), it is relatively short ( 17 mm, i.e. 33% of the length of the cheek teeth row LRDJ [ LP2/-M3/], Appendix 20A), but on AUM 1544, it could have exceeded 50% of LRDJ. On mandibles, the length of the DPC can only be known indirectly on AUM 161, which preserves the molar row but is crushed in the premolar region. With a molar length of 33.6 mm and assuming a PMI between 0.67 and 0.70 (an average value for a Pachynolophus ), the LRDJ length can be estimated at 56-57 mm. Furthermore, the socket of the canine is missing and the part of the DPC preserved until the anterior alveolus of P/2 is 23 mm long. The DPC would not therefore have been lower than 40 or 41% of LRDJ. With a length of 33 mm, the DPC of AUM 167 would have had a greater relative value, probably over 50% (Appendix 20B).
Pachynolophus ruscassierensis n. sp.
P2/ P3/ P4/ M1/ M2/ M3/
L W L W L W L W D d L W D d L W D d
N 2 2 7 7 8 8 18 18 17 19 18 19 18 18 11 11 11 11 mean 6.3 5.7 7.8 9.4 8.3 10.4 9.8 11.8 12.5 11.6 10.9 12.9 13.7 12.5 11.1 13.0 13.8 12.7 variation 6.0- 5.5- 7.1- 9.1- 7.3- 9.4- 9.0- 10.2- 10.8- 10.3- 9.2- 10.1- 11.1- 10.1- 10.3- 12.1- 12.6- 11.4- range 6.5 5.8 8.5 9.9 9.2 11.6 10.7 13.2 14.6 12.9 12.2 14.7 15.9 14.2 12.1 14.3 15.4 13.9 standard – – 0.549 0.263 0.657 0.856 0.554 0.766 0.995 0.871 0.939 1.292 1.374 1.371 0.662 0.737 0.962 0.885 deviation
coeff. of – – 7.0 2.8 8.0 8.3 5.7 6.5 8.0 7.5 8.6 10.0 10.0 11.0 5.9 5.7 6.9 7.0 variation
DP2/ DP3/ DP4/
L WD d L WD d L WD d
N 1 1 1 1 2 2 2 2 1 1 1 1
mean 7.4 5.9 8.7 5.8 9.0 10.0 10.5 9.8 9.5 11.2 11.9 11.5
variation range – – – – 8.9- 9.8- 10.4- 9.7- – – – –
9.0 10.1 10.6 9.8
upper series LDPC LP2-M3 %DPC LP2-P4 LM1-M3 PMI
N 2 2 2 4 3 3
mean 22.3 52.2 42.5 21.8 31.9 68.4
variation range 17.0-27.5 51.3-53.1 33.1-51.8 20.5-22.6 31.2-32.4 65.7-69.4
P/2 P/3 P/4 M/1 M/2 M/3
L W1 W2 L W1 W2 L W1 W2 L W1 W2 L W1 W2 L W1 W2 W3 N 3 3 3 3 3 3 5 5 5 12 12 11 11 11 11 7 9 8 7 mean 7.3 3.8 4.1 8.1 4.6 5.4 9.0 5.7 6.0 9.1 5.9 6.2 10.7 7.0 7.0 14.2 6.8 6.3 4.9 variation 6.7- 3.7- 3.8- 7.2- 4.3- 5.2- 8.3- 5.1- 5.5- 8.3- 5.4- 5.9- 9.4- 6.1- 6.5- 13.4- 6.2- 5.6- 4.3- range 7.6 3.9 4.3 8.7 4.7 5.8 9.3 6.0 6.2 10.2 6.5 6.8 12.4 8.1 7.8 15.2 7.7 7.1 5.9 standard 0.520 0.100 0.265 0.814 0.231 0.321 0.412 0.356 0.277 0.550 0.291 0.401 0.927 0.625 0.395 0.655 0.559 0.677 0.568 deviation
coeff. of 7.1 2.6 6.5 10.0 5.1 5.9 4.6 6.2 4.6 6.0 4.9 6.5 8.7 8.9 5.6 4.6 8.3 10.8 11.5 variation
DP/2 DP/3 DP/4
L W1 W2 L W1 W2 L W1 W2
N – – – 1 1 1 3 4 3
mean – – – 7.9 4.5 5.7 8.5 5.4 5.6
variation range – – – – – – 8.3-8.7 5.1-6.0 5.4-5.9
standard
deviation – – – – – – 0.208 0.403 0.252
coeff. of variation – – – – – – 2.5 7.4 4.5
lower series LDPC LP2-M3 %DPC LP2-P4 LM1-M3 PMI
N 1 1 1 1 2 1
mean> 23.0 56.7> 41.0 22.5 34.2 67.0
variation range – – – – 33.6-34.8 –
In the Aumelas sample neither the deepness of the nasal notch on the skull, nor the shape of the premaxillary vertical apophysis are known. The FIO is rather rostrally situated, at 7 mm above P3/ (AUM 231, AUM 1552). On the juvenile AUM 1618, it appears above DP2/. The anterior edge of the orbit (O) levels the mesial border of M2/. The orbital floor is very low (HTMX about 5 mm), and moreover hilly by the relief of the roots of the last molars. The palate is slightly crushed (AUM 231) but appears hollow. It seems neverthe- less have been rather narrow after correction of that crash (taking into account the position of the spine of the median raphe). The maxillary zygomatic process, which is not very dorso-ventrally high ( 7 mm at orbit level) is expanded laterally, but the bow does not seem to have strongly diverged at the back, like on Pachynolophus livinierensis .
On AUM 161, the mandibular body is not very high and only increases slightly backwards ( 13.3 mm under DPC; 16.8 mm under P/2; 23.4 mm under M/3). The values are close for AUM 169. On the juveniles AUM 1556 and AUM 1605, the mandibular height increases from 16 mm under DP/ 3 to 18 mm under M/2. Two mental foramina are noticeable, one under the limit P/3-P/4 and the second under P/1-P/2 (AUM 161). The vertical ramus is not preserved in Aumelas sample.
COMMENTS
The described material of Aumelas can only be assigned to a Pachynolophus , based on the lack of mesostyle on upper molars, on the weak molarization of the premolars, especially with the upper ones devoid of hypocone. This form is first larger than most Pachynolophus species. Only one, Pa. cayluxi, known only by two upper premolars, P3/ and P4/, is markedly larger. The P4/ of Pa. cayluxi is also more transversely elongated with a narrower lingual contour.
On the contrary, the cheek teeth of the lower Eocene species Pa. eulaliensis are significantly smaller, as well as those of other potentially comparable Equoidea, Hyracotherium remyi or Orolophus maldani (on average 28 to 35% smaller than Aumelas’ form). So great are these differences that it seems useless to present comparative tables of measures. Moreover, these species are on the whole more bunodont and slightly more brachyodont; the cingula are wider and higher. The parastyle of molars is more overflowing in H. remyi. The P4/ of Pa. eulaliensis has a mesially shifted protocone and an accessory cusp lingual to the metacone.
The Middle Eocene genotypic species Pa. duvali is also smaller than the species of Aumelas with differences around 20% for most parameters. The ectoloph of upper cheek teeth is less flat in Pa. duvali . P4/ and P3/ are lingually narrower, their lingual cingulum is weaker.According to the mandible MNHN CGR-89, the DPC should have been much longer. Then, Pa. livinierensis is about 25% smaller than the Aumelas’ species with a smaller premolar sector and a proportionally greater surface ratio M3/ on M1/ (Appendix 21). The cingula are also weaker in Pa. livinierensis and the DPC longer. Pachynolophus gaytei, which is very small, has more transversely elongated and narrower premolars.
Two Spanish species have roughly the same size as that of Aumelas: Pachynolophus molipontiensis and Pa. boixedatensis . The former is characterized (Checa-Soler 1994, 1997: pl. 2 fig. 7) by its low premolar surface, with a very small SP / SM ratio of only 34% instead of 44-49 in the sample from Aumelas (Appendix 21). The second, which is however a bit smaller than the Aumelas’ species, distinguishes chiefly by the less transversely elongated P4/, with a more lophodont postprotocrista devoid of a conspicuous metaconule. Moreover, in Pa. boixedatensis , the DPC is likely longer (ICP 3368), the upper molars have an internal outline less recessed labially at the level of the hypocone, and the M3/ is proportionally larger.
The species Pa. cesserasicus , from the Minervois area, could present some affinities with the material from Aumelas but it is too badly defined and needs more extensive descriptions and complete revision ( Savage et al. 1965) before concluding. Assuming that the featured specimen FSL-2977 could be a good representative of Pa. cesserasicus and that both parts of this specimen pertain to the same individual, we can observe that its size falls within the range of the sample from Aumelas. Besides, the lingual cingulum of premolars is almost continuous in both species. But significant differences are also observed and rules out assignment of the material of Aumelas to this taxon. Indeed, in Pa. cesserasicus the dentition seems more bunodont with more bulged cusps and less flattened ectolophs. The premolar area is smaller ( SP / SM 39%; Appendix 21). P4/ is more transversely elongated and narrower on lingual side. The labial cusps of P3/ are close to each other. Furthermore, Pa. cesserasicus has an extremely long DPC, the longest observed in any Pachynolophus .
Special attention must be brought to the Rouzilhac taxon attributed by Godinot et al. (2018) to “ Propachynolophus ” maldani (synonym of Orolophus maldani, see Remy 2017), which in fact has a size close to Pachynolophus ruscassierensis n. sp. according to the measures mentioned by the authors and from the published pictures.
The Rouzilhac material is much larger than the species of the Paris Basin. Indeed the length of M/3 varies between 13.5 and 15.7 mm at Rouzilhac ( Godinot et al. 2018: table 12), vs a length of 10.9 mm for the holotype of O. maldani (MNHN AL-5199), and the length M/1-M/3 from 31.3 to 34.9 mm vs 23.5 mm (Appendix 25). Similar differences occur with upper cheek teeth. For most parameters, the sample of Rouzilhac is +21% larger on average than the material of the species maldani, a highly significant difference (p = 0.001).
Conversely molars of Pachynolophus ruscassierensis n. sp. are rather similar in size to the Rouzilhac sample (at Aumelas, LM/3 13.4-15.2 mm; LM/1-M/3 33.6-34.8 mm). Both samples share several anatomical features (on the skull, location of orbit and of FIO and level of choanae; at the mandible, length of DPC).
It might therefore be asked whether the Rouzilhac sample should be referred to the Aumelas species. Nevertheless, a large amount of arguments invalidates such a hypothesis.
First, whereas molars have almost similar size, the premolars are smaller at Rouzilhac (particularly P3/ and P/4). Thus in surface the premolar sector is smaller, revealing an unlike distribution of masticatory function ( SP / SM 39 on the upper series RZ-009, vs 44-49 at Aumelas [Appendix 21]; SP 2-P3/3M 19 vs 25-28). Same differences occur with lower cheek teeth. Premolars are on average 12% smaller for most parameters (–26% in area), these differences concerning above all the P/4 (significant up to level p = 0.001).
Besides, the upper molars of the Rouzilhac sample are more trapezoidal in shape, more bunodont, with not so flat ectolophs. In addition some bear a mesostyle, sometimes well defined ( Godinot et al. 2018: 266), whereas this feature is seemingly totally absent in Pa. ruscassierensis n. sp. The lower premolar ectocingulid is more pronounced at Rouzilhac. Finally the mandibular symphysis is lightly more elongated (ending at the level of P/1 instead of 5 mm further forward).
Thus, it seems that the Rouzilhac sample assigned by Godinot et al. to “ Propachynolophus ” maldani cannot be confused with Pachynolophus ruscassierensis n. sp. from Aumelas, and that it likely represents an original taxon also differing from Orolophus maldani of the Paris Basin.
Concerning the youngest Pachynolophus species, Pa. garimondi and Pa. bretovensis differ by thinner and lower cingula, by less deeply hollowed centrocristae on upper cheek teeth, and by a wider distal basin on P4/. Pachynolophus lavocati and Pa. zambranensis are distinguished by a more marked lophodonty than in specimens of Aumelas, including fully fused conules in transverse lophs, and Pa. lavocati presents a reduced premolar sector ( SP / SM = 37%) (Appendix 21).
The species from Aumelas finally represents an original form that cannot be reported to any other known species of the genus. In the context of evolutive trends observed in Pachynolophus (e.g. Remy 2017: fig. 6, 18, supp. data 4, 12), this new species that we name Pa. ruscassierensis n. sp., combines likely archaic features (persistence of relatively thick and high cingula, not greatly reduced premolar sector, relatively short DPC) and some ones seemingly more derived (fairly flat ectoloph of upper molars, rather wide internal outline of the P/).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Pachynolophinae |
|
Genus |