
Pristimantis muchimuk Barrio-Amorós, Mesa, Brewer-Carías & McDiarmid, 2010
|
publication ID |
https://doi.org/10.11646/zootaxa.3686.3.3 |
|
publication LSID |
lsid:zoobank.org:pub:F503F48B-57B7-498F-91B4-C975DAA6956C |
|
DOI |
https://doi.org/10.5281/zenodo.5614495 |
|
persistent identifier |
https://treatment.plazi.org/id/038487A0-9738-4012-FF0E-F128D6CA55A2 |
|
treatment provided by |
Plazi |
|
scientific name |
Pristimantis muchimuk Barrio-Amorós, Mesa, Brewer-Carías & McDiarmid, 2010 |
| status |
|
Pristimantis muchimuk Barrio-Amorós, Mesa, Brewer-Carías & McDiarmid, 2010
( Figures 6–7 View FIGURE 6 View FIGURE 7 , Tables 1–2 View TABLE 1 View TABLE 2 )
Material examined. A female ( MHNLS 19652, Holotype), from a step of the northern face of Churí-tepui (base camp of the Muchimuk-Expedition 2009), Macizo de Chimantá , Parque Nacional Canaima, Gran Sabana municipality, Bolívar state, Venezuela ( 05°16'45"N, 62°00'56"W; elevation 2325 m), collected on May 2009, by I. Elorza, J. Mesa and C. Brewer-Carías; three males ( MHNLS 20495–97, field numbers PS 342–44, respectively) one female ( MHNLS 20505, field number PS 353) and a juvenile ( MHNLS 20506, field number PS 354), from Churí-tepui, Macizo de Chimantá , Parque Nacional Canaima, Gran Sabana municipality, Bolívar state, Venezuela ( 05°15'16.6"N, 62°00'31.6"W; elevation 2383 m), collected on July 29–30, 2011, by F.J.M. Rojas-Runjaic, P.E. Salerno and G.B. Pauly.
Remarks. Barrio-Amorós et al. (2010) described Pristimantis muchimuk based only on a single female ( holotype MHNLS 19652) which was poorly preserved and dehydrated. The ventral skin was described as smooth; however, observations of this character, both in life and in preservative, based on the five new specimens, show that the ventral skin is areolate (or coarsely areolate) rather than smooth, and that the smooth aspect of the ventral skin in the holotype is an artifact of its poor state of preservation. Additionally, Barrio-Amorós et al. (2010) did not mention the color pattern of the axillae, groins and hidden parts of the hind limbs because the description of color in life was based on a single photograph of the holotype in profile ( Fig. 6 View FIGURE 6 a). The inspection of the new series of Pristimantis muchimuk indicates that all specimens (three adult males, one female, and one juvenile) have a bright red coloration in these regions ( Figs. 6 View FIGURE 6 b–d, 7a–d). This coloration may be a fixed character in P. m u c h i m u k and may be useful for identification because it is not common in other Pristimantis of the Guiana Shield region. Based on the above mentioned differences, and observations of additional diagnostic characters in the males from the new series, we propose an amended diagnosis for this species.
Diagnosis amended. (1) body size small, with adult females ranging from 20.5–25.2 mm SVL, and adult males from 18.1–19.4 mm SVL; (2) dorsal skin shagreen to warty, warts smooth, low and flat, with a barely distinguishable middorsal raphe; (3) ventral skin areolate to coarsely areolate; (4) tympanic annulus and membrane absent; (5) snout rounded in dorsal view, rounded to nearly truncate in profile; (6) canthus rostralis nearly straight in dorsal view; rounded in cross section; (7) upper eyelid smooth, with one barely discernible tubercle; (8) choanae small, oval; (9) dentigerous processes transverse to diagonal, small, posterior and medial to choanae, each bearing one to five teeth; (10) tongue rounded posteriorly; (11) males without vocal slits or nuptial pads; (12) FI shorter than FII; (13) fingers with lateral keels; (14) ulnar tubercles absent; (15) tarsal tubercles and calcars absent; (16) tarsal fold absent (17) inner metatarsal tubercle oval, three times the size of the round outer metatarsal tubercle; (18) toes with lateral fringes; (19) basal webbing between TIII–TV; (20) TV longer than TIII; (21) axillae and groins colored bright red in life.
Variation. Pristimantis muchimuk does not have gender-specific coloration, and is quite variable in color pattern in both sexes ( Fig. 6 View FIGURE 6 a–f). Additionally, males lack vocal slits and nuptial pads, making it very difficult to differentiate males and females at first glance. Except for size (females larger than males) no other sexually dimorphic characters are noticeable. Males and females of similar size could only be identified by dissection and gonad examination. Adult females range from 20.5–25.2 ( n = 2), and adult males from 18.1–19.4 ( n = 3). Variation of morphometric characters is shown in Table 1 View TABLE 1 . Head length ranges from 39.2–41.1% SVL ( n = 5), and maximum head width from 37.6–38.6% SVL ( n = 5). The ratio EN/ED is slightly variable but every eye-naris distance is smaller than eye diameter (0.8–1.0 [ n = 5]). Also, the ratio UEW/IO is somewhat variable in males (0.7–0.9 [ n = 3]), but variation in this ratio is not appreciable in females (UEW/IO = 0.7; n = 2). The snout is slightly shorter and more variable in males than females (males: 1.1–1.3 [ n = 3], vs. females: 1.2–1.3 [ n = 2]), but always larger than eye diameter. Hand length ranges from 24.6–27.0% SVL ( n = 5) with hands slightly larger in females than males (females: 26.1–27.0% SVL [ n = 2], vs. males: 24.6–25.2% SVL [ n = 3]). Shank length ranges from 48.6–50.9% SVL ( n = 5) but in this case, males have shanks slightly longer than females (males: 50.1–50.9% SVL [ n = 3], vs. females: 48.6–49.2% [ n = 2]). Variation in other morphometric ratios is not relevant.
Ventral skin of belly coarsely areolate in MHNLS 20495 and 20497 (males), areolate in MHNLS 20496 (male) and 20505 (female); slightly areolate in MHNLS 20506 (juvenile). Snout rounded in dorsal view and profile in MHNLS 20495–20497 and 20505, subacuminate in dorsal view in the juvenile MHNLS 20506 (nearly truncate in profile in the holotype). Tubercle on upper eyelid absent in MHNLS 20505, barely visible in all other specimens in life (not visible in preservative). Dentigerous processes and number of teeth somewhat variable: diagonal and very low dentigerous processes, with one to two teeth in MHNLS 20497; diagonal, prominent and also with two to three teeth in MHNLS 20496; transverse, prominent and with two to three teeth in MHNLS 20495; almost transverse and prominent, with three to four teeth in MHNLS 20505. One ulnar tubercle barely visible (near the hand) in MHNLS 20496 and 20497, not visible in the other specimens examined. Bright red coloration on axillae, groins and hidden parts of the hind limbs variable in extent and intensity ( Figs. 7 View FIGURE 7 a–d), but present in the five new specimens collected. No additional variation was observed in other discrete morphological characters.
Advertisement call. Calls of Pristimantis muchimuk were recorded on the night of June 30th 2011 between 2200 and 2300 h at an air temperature of approximately 13°C. We recorded advertisement calls of two males that were calling at the border of a Bonnetia roraimae forest adjacent to a creek (Churí-tepui, 0 5° 15.257’N, 62° 00.472’ W). The two males were calling in close proximity, and several calls by individual 1 needed to be excluded from the analyses because they overlapped with calls of individual 2. As a result, we analyzed 11 calls from individual 1 and 25 from individual 2. A summary of the call parameters for each male and for both averaged can be found in Table 2 View TABLE 2 . We could not be certain that the recorded males were in fact the males collected because the calling individuals were hidden inside Brocchinia hechtioides tubes ( Bromeliaceae ) and within rosettes of Orectanthe sp. ( Xyridaceae ). Nonetheless, no other species of frog was found at that spot, the males collected were adult, and were found in the sites from which the calls emanated.
Low Freq. (Hz) High Freq. (Hz) Call length (s) Dom. Freq. (Hz) Bandwidth (Hz)
X? ± SD X? ± SD X? ± SD X? ± SD X? ± SD The calls of Pristimantis muchimuk consist of a single soft note that resembles two pieces of glass lightly bumping each other. Calls are performed in bouts, and the longest bout recorded had a total of 25 calls. The call rate was 2.07 calls/sec for individual 1 and 1.72 calls/sec for individual 2, counting only within-bout time. For individual 2, inter-call intervals decreased throughout the bout, with an average inter-call interval of 0.95 s at the beginning of the bout (first five calls) and 0.45 s at the end of the call bout (last five calls). The average call duration was 0.041 s (range: 0.027– 0.062 s). The average dominant (fundamental) frequency was 2684.45 Hz with a frequency range between 2180–3399 Hz ( Fig. 8 View FIGURE 8 ).
Comparisons with calls of other Guiana Shield highland Pristimantis . Of the six highland Pristimantis with described advertisement calls, Pristimantis muchimuk has a similar call structure to P. aureoventris of Wei Assiputepui and Mount Roraima (Kok et al. 2011), P. pruinatus of Cerro Yaví ( type 2 call of Myers & Donnelly 1996), and P. yuruaniensis of Yuruaní-tepui ( Rödder & Jungfer 2008), in that they all produce bouts of single note calls of similar duration and call rate. Note, that what we and others ( Myers & Donnelly 1996, Kok et al. 2011) have considered a call bout consisting of multiple calls, others ( Myers & Donnelly 1997, Rödder & Jungfer 2008) termed a single call consisting of multiple pulses. In terms of spectral structure, the most similar call is P. pruinatus in that it is the only other vocalization that lacks visible harmonics, has a similar dominant frequency (2440–2660 Hz), and lacks frequency modulation ( Myers & Donnelly 1996). However, inter-call interval is shorter in P. muchimuk (0.397– 2.199 s) than in P. pruinatus (1.5– 2.9 s; Myers & Donnelly 1996), and more similar to P. yuruaniensis (0.504– 1.968 s; Rödder & Jungfer 2008). Pristimantis yuruaniensis has much lower dominant frequency (1.86–2.08), with two visible harmonics ( Rödder & Jungfer 2008). Pristimantis aureoventris has a much longer inter-call interval (3.097 ± 0.275) as well as obvious harmonics and frequency modulation of the first note (Kok et al. 2011). The remaining two highland species differ from P. m u c h i m u k in that P. cantitans has a longer inter-call interval of 2.3– 7.8 s ( Myers & Donnelly 1996), and P. memorans has calls which were described as loud “tinks” in groups of 1–4 ( Myers & Donnelly 1997).
Distribution. Pristimantis muchimuk is known only from two locations, both on the summit of Churí-tepui (Barrio-Amorós et al. 2010; this work), and separated by a distance of ca. 2.8 km ( Fig. 1 View FIGURE 1 ). The altitudinal range is 2325–2383 m. Churí-tepui is one of the 11 mountains that makes up the Chimantá massif and is located in the southern section of the massif, reaches a maximum elevation of ca. 2500 m and has a summit area of 47.5 km 2 ( Huber 1995).
Habitat and natural history. The five new specimens of P. m u c h i m u k were found at night within a 10 m radius; all were a few meters from a small seasonal creek, found in vegetation 0.2 to 1.2 m above the ground, and within a small grove of Bonnetia roraimae ( Figs. 9 View FIGURE 9 a–b). The first three individuals, all adult males, were collected the evening of June 29, 2011 between 2100 and 2115 h. There had been periodic rain throughout the day. The presence of the frogs was detected by their chorusing activity. The first male (MHNLS 20495) was collected from a green side leaf of a Brocchinia hechtioides ; this individual was perched on the upper surface of the leaf and retreated further back into the leaf axil upon disturbance. The second male (MHNLS 20496) was also collected from a Brocchinia hechtioides but this time from within the central tube of the plant. The third male (MHNLS 20497) was found within the green leaves that make up the rosette of an Orectanthe sp. The following evening, after no rain since the previous night, chorusing activity was again detected from the same Bonnetia roraimae grove, although the chorusing activity was less intense than the previous evening. Nevertheless, calls from two males were recorded. Furthermore, two individuals were found including a female (MHNLS 20505) and a juvenile (MHNLS 20506); both were found in side leaves of Brocchinia hechtioides . The female ( 20.5 mm SVL) was smaller than the female holotype ( 25.2 mm SVL) described by Barrio-Amorós et al. (2010), but the presence of large oviducal eggs discovered via dissection indicated that it was an adult. Although the chorus was loud enough to be heard from approximately 20 m away, no other calls were detected in the area outside of the small Bonnetia grove. Interestingly, there were no other close areas with both Bonnetia trees and a flowing creek, thus suggesting that the two components together may be indicative of a breeding habitat.
Although there are few available observations of Pristimantis species that inhabit high-elevation tepui summits, some patterns seem to be emerging. First, most species, including P. abakapa , P. aureoventris , P. marahuaka , P. m u c h i m u k, P. pruinatus , and P. yaviensis , seem to be largely nocturnal and associated with vegetation (Barrio-Amorós et al. 2010, Kok et al. 2011, Myers & Donnelly 1996, Fuentes-Ramos & Barrio- Amorós 2004). The exceptions include P. cantitans , which calls during the day and night and was found in vegetation and also below moss mats over sandstone ( Myers & Donnelly 1996) and P. yuruaniensis which, at least in captivity, showed diurnal as well as nocturnal activity ( Rödder & Jungfer 2008). Second, although specific plant associations have only been described for P. abakapa , P. aureoventris , P. marahuaka and P. m u c h i m u k (Fuentes- Ramos & Barrio-Amorós 2004, Barrio-Amorós et al. 2010, Kok et al. 2011, and this work), there appears to be close associations with Brocchinia sp. for all four species, with Orectanthe sp. for P. m u c h i m u k and P. aureoventris , and with Heliamphora sp. for P. marahuaka .



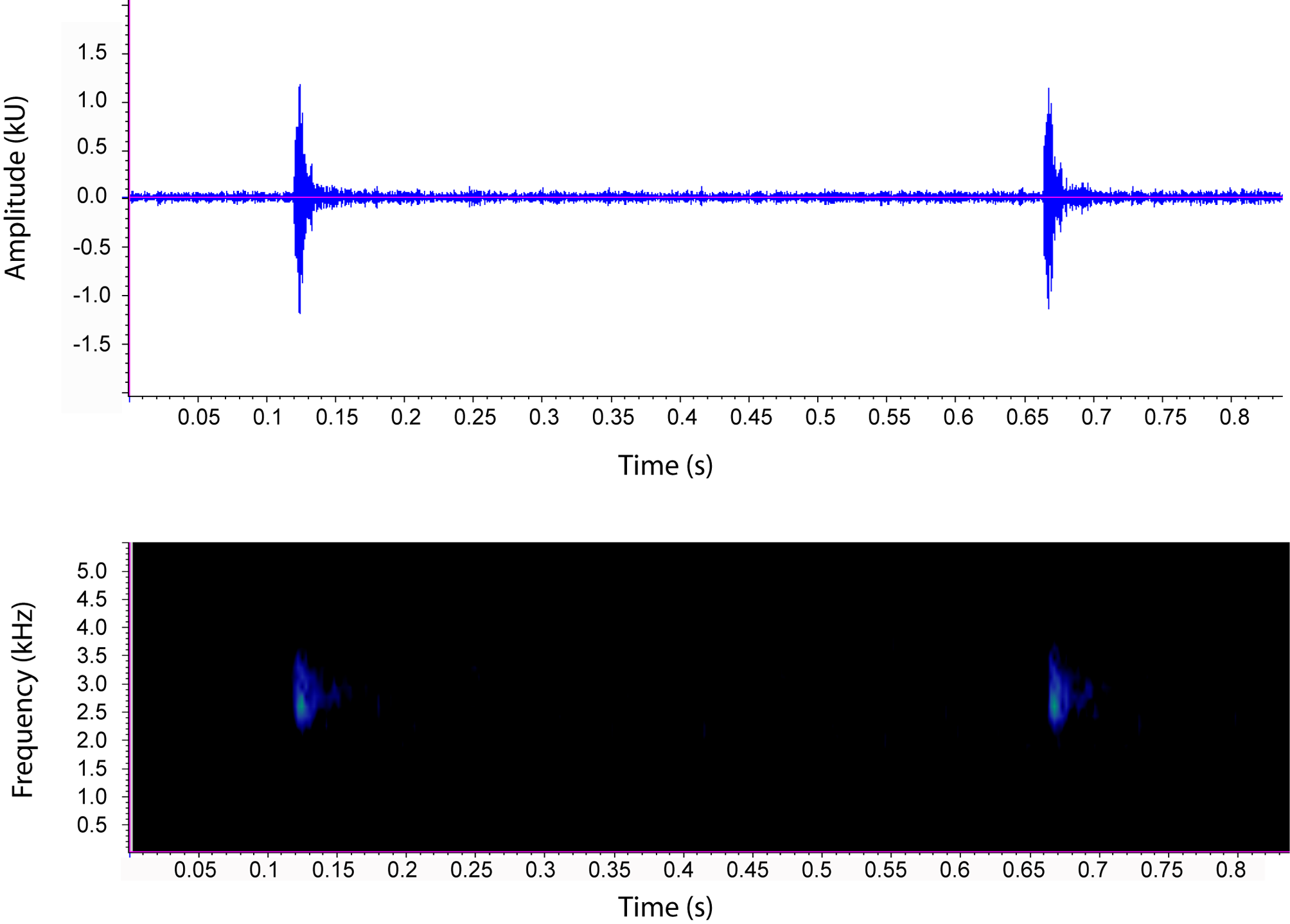

TABLE 1. Morphometric variation (in mm) in the five adult specimens (three males and two females) of Pristimantis muchimuk. X? ± SD: mean ± standard deviation; Min – Max: minimum – maximum.
| Morphometric characters Males ( n = 3) | Females ( n = 2) | ||
|---|---|---|---|
| X? ± SD | Min–Max | X? ± SD | Min–Max |
| SVL 18.5 ± 0.8 | 18.1 – 19.4 | 22.9 ± 3.3 | 20.5 – 25.2 |
| ThL 8.7 ± 0.5 | 8.2 – 9.2 | 11.0 ± 1.9 | 9.7 – 12.4 |
| SL 9.3 ± 0.3 | 9.1 – 9.7 | 11.2 ± 1.7 | 10.0 – 12.4 |
| FL 7.2 ± 0.5 | 6.7 – 7.6 | 8.9 ± 1.0 | 8.2 – 9.6 |
| HaL 4.6 ± 0.2 | 4.5 – 4.8 | 6.1 ± 1.0 | 5.4 – 6.8 |
| HeL 7.5 ± 0.4 | 7.1 – 7.9 | 9.1 ± 1.1 | 8.3 – 9.9 |
| HW 7.0 ± 0.2 | 6.9 – 7.3 | 8.8 ± 1.2 | 7.9 – 9.6 |
| IO 2.3 ± 0.0 | 2.3 – 2.4 | 2.8 ± 0.1 | 2.7 – 2.9 |
| UEW 1.9 ± 0.2 | 1.7 – 2.1 | 2.0 ± 0.1 | 1.9 – 2.1 |
| IND 1.8 ± 0.1 | 1.7 – 2.0 | 2.1 ± 0.1 | 2.1 – 2.2 |
| EN 2.2 ± 0.1 | 2.1 – 2.2 | 2.7 ± 0.4 | 2.4 – 3.0 |
| ED 2.5 ± 0.2 | 2.3 – 2.8 | 3.0 ± 0.2 | 2.8 – 3.2 |
| TD – | – | – | – |
| ETS 3.0 ± 0.1 | 2.9 – 3.1 | 3.8 ± 0.5 | 3.4 – 4.2 |
| F3D 0.9 ± 0.0 | 0.9 – 1.0 | 1.0 ± 0.0 | 1.0 – 1.0 |
| T4D 0.9 ± 0.0 | 0.9 – 0.9 | 0.9 ± 0.0 | 0.9 – 0.9 |
| Fi1L 2.9 ± 0.3 | 2.6 – 3.2 | 3.7 ± 0.5 | 3.4 – 4.1 |
| Fi2L 3.4 ± 0.2 | 3.3 – 3.7 | 4.3 ± 0.5 | 4.0 – 4.7 |
TABLE 2. Advertisement call characteristics for two males of P. m u c h i m u k. The values are shown for each specimen separately, and for the average.
| Individual 1 (11 calls) | 2352.85 ± 41.38 3081.75 ± 63.48 | 0.039 ± 0.012 | 2709.24 ± 80.43 | 728.0 76.73 |
|---|---|---|---|---|
| Individual 2 (25 calls) | 2103.95 ± 99.2 3539.31 ± 155.2 | 0.043 ± 0.005 | 2673.55 ± 112.5 | 1435.35 247.26 |
| X? (2 indiv.) | 2228.40 3310.53 | 0.041 | 2691.40 | 1081.67 |
| MHNLS |
Coleccion de Mastozoologia, Museo de Historia Natural de La Salle |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |