
Choerophryne gracilirostris, Richards, Amy Iannella Stephen & Oliver, Paul, 2014
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3753.5.6 |
|
publication LSID |
lsid:zoobank.org:pub:93D3FEB4-3C66-4518-AA4A-67F2B091F46A |
|
DOI |
https://doi.org/10.5281/zenodo.5627913 |
|
persistent identifier |
https://treatment.plazi.org/id/03856B5A-FFCB-FFF0-FF29-FA051C647253 |
|
treatment provided by |
Plazi |
|
scientific name |
Choerophryne gracilirostris |
| status |
sp. nov. |
Choerophryne gracilirostris View in CoL sp. nov.
( Figs 1–5 View FIGURE 1. A View FIGURE 2 View FIGURE 3 View FIGURE 5 )
Holotype. SAMA R67520 (Field No. SJR 10506), adult male, calling when collected, upper Strickland River catchment, Western Province, Papua New Guinea (5.8078o S, 142.3083o E, 213 m a.s.l.), collected by S. Richards 29 March 2008.
Paratopotype. SAMA R67519 (Field No. SJR 10505) adult male, with same collection details as holotype.
Paratypes. SAMA R67514 (Field No. SJR 10230), SAMA R67516 (Field No. SJR 10267), SAMA R67517 (Field No. SJR 10269), Karius Range, Southern Highlands Province, Papua New Guinea (5.9911o S, 142.6706o E, 1368 m a.s.l.), collected by S. Richards 6–8 February 2008, SAMA R67518 (Field No. SJR 10420) upper Strickland River catchment, Western Province, Papua New Guinea (5.9018o S, 142.6706o E, 953 m a.s.l.), Western Province, Papua New Guinea, collected by S. Richards 20 February 2008, SAMA R64986 (Field No. SJR 10946). Gugusu Camp, Muller Range, Western Province, Papua New Guinea (5.7291o S, 142.2632o E, 515 m a.s.l.), 9 September 2009.
Referred material. SAMA R67515 (Field No. SJR 10234) Juvenile, Karius Range, Southern Highlands Province, Papua New Guinea (5.9911o S, 142.6706o E, 1368 m a.s.l.), collected by S. Richards 6 February 2008.
Diagnosis. Assigned to the genus Choerophryne on the basis of its distinctly projecting snout, comprising an extension of the nasal bones and the alary processes of the premaxillae, and the posterolateral expansion of the sacrum, fused with the urostyle (as confirmed by CT scanning). A small Choerophryne (SUL 13.5–14.7 mm) with a distinctly projecting, elongated and narrow snout (OHG/SUL 0.06–0.08), urostyle projecting posteriorly beyond the insertion of the hind limbs, leg length average for genus (TL/SUL 0.37–0.40), expanded discs on all digits of hands and feet except the first, but not more than 1.4 times width of penultimate phalanges, distinctive dark-brown hourglass marking on the dorsum, and advertisement call consisting of 3–5 distinctly pulsed notes repeated at a rate of 0.36–0.38/s for up to one minute.
Comparison with other species. Choerophryne gracilirostris sp. nov. is distinguished from the two congeners known from the southern versant of the central cordillera ( C. allisoni and C. burtoni ) by its larger size (SUL 13.5–14.7 mm vs. 11.5–11.6 mm in C. allisoni and 12.1–13.6 mm in C. burtoni ), and much longer snout (OHG/SUL = 0.06–0.09 vs. 0.03–0.06 in both species); it further differs from C. burtoni in being considerably less tuberculate, especially towards the anterior portion of the dorsum and in having calls with much shorter notes (0.014– 0.042 s vs 0.18– 0.29 s) and fewer pulses (4–9 vs 9–15).
Of the geographically disjunct (northern) species Choerophryne gracilirostris sp. nov. is readily distinguished from C. longirostris , C. microps , C. nigrescens and C. proboscidea by its smaller size (SUL<14.7 mm vs.> 15.2 mm); all of these taxa except C. microps also differ in having a distinctly widened disc on the first finger. Choerophryne gracilirostris sp. nov. further differs from C. longirostris and C. microps in having a relatively shorter snout (OHG/SUL = 0.06–0.09 vs. 0.09–0.11) and from C. nigrescens in having a light buff venter with light brown vermiculations (vs. darker blackish brown vermiculations) and in having (vs. lacking) a dorsal hourglass marking.
Three further Choerophryne are all of similar size to Choerophryne gracilirostris sp. nov. (13.5–14.7 mm). The new species can be distinguished from C. amomani by its longer snout (SL/SUL 0.21–0.26 vs 0.18–0.21), longer tibiae (TL/SUL 0.37–0.40 vs. 0.33–0.38) and call with 3–5 notes (vs 1–2); from C. rostellifer by its slightly smaller size (male SUL 13.5–14.7 mm vs. 14.0– 16.8 mm (18.0 mm for female C. rostellifer ), generally smaller disks on the third finger (F3D/SUL 0.03–0.04 vs. 0.03–0.06 in C. rostellifer ) and notes with 4–9 (vs ~14) pulses. The snout of C. gracilirostris sp. nov. is also more slender than in C. rostellifer , tapering more rapidly to achieve a narrower tip in dorsal aspect ( IND /SUL 0.05–0.06 vs. 0.06–0.09). Despite being only distantly related (A. Iannella, unpublished data), in size and appearance Choerophryne gracilirostris sp. nov. is most similar to C. arndtorum . It is on average slightly larger (SUL 13.5–14.7 mm vs. 11.2–14.8 mm), tends to have a slightly longer snout (SL/SUL = 0.21–0.26 vs. 0.19–0.22) and has a distinctly different call comprising notes lasting just 0.014– 0.042 s (vs 0.13– 0.18 s in C. arndtorum ) and containing just 4–9 pulses (vs 22–44 in C. arndtorum ).
Description of holotype. Adult male with a short ventral incision on right side. Head moderately wide (HW/ SUL 0.35), canthus rostralis weakly defined, loreal region very slightly concave. Snout narrow, acute and projecting well beyond anterior rictus of lower jaw (OHG 1.2 mm), tip rounded in dorsal and lateral profiles, ventral surface of tip with a broad, unpigmented medial ridge, widest anteriorly and tapering towards anterior rictus of the upper jaw. Nares oval, narrowing slightly posteriorly, located close to tip of snout, oriented ventrolaterally, not visible in dorsal view, internarial distance small (0.8 mm). Eye moderately small (ED/SUL 0.09), transverse diameter less than length of snout, pupil horizontal. Tympanic annulus indistinct, ventral edge faintly apparent. Tongue oval, two curved prepharyngeal ridges present, vomerine teeth absent, vocal slits present slightly anteriolateral to base of tongue. Urostyle projecting beyond vent, giving posterior of torso acute appearance in both dorsal and ventral aspects. Dorsum finely granular with faint medial ridge extending from snout tip to urostyle tip and scattered small low and indistinct tubercles across flanks and dorsal surface of hindlimbs; venter smooth.
Fingers moderately long and slender with distinct circum-marginal grooves, relative lengths III>IV>II>I, finger I less than half length of II, fingers II–IV with distinctly expanded discs up to 1.3 times width of penultimate phalanx, finger I without disk and with only faint circum-marginal groove; subarticular tubercles and webbing absent. Legs of moderate length (TL/SUL 0.40). Toes moderately long and wide, relative lengths IV>III>V>II>I, toe I approximately half length of II, all toes except I with terminal discs up to 1.4 times width of penultimate phalanx and with circum-marginal grooves; metatarsal tubercles and webbing absent. Disks on toes wider than those on fingers (F3/T4 0.78, Fig. 1 View FIGURE 1. A ).
In preservative, ground colour of dorsum medium brown with dense dark-brown maculations, slightly denser anteriorly than posteriorly; dorsum dominated by a longitudinally oriented dark-brown hourglass mark, defined laterally by a pair of pale-tan dorso-lateral triangles extending from dorsal edge of the tympanum to the groin with medial tips approaching mid-dorsum, these pale-tan triangular patches with prominent but weakly defined darkbrown edging. Inguinal region with a pair of indistinct dark-brown lumbar ocelli with very small, distinct, pale cream medial spots. Vent bordered laterally and dorsally by an arc of dark-brown pigment. Exposed dorsal and lateral surfaces of limbs pale tan, extensively flecked and blotched with various shades of darker brown; darker pigmentation on limbs tending not to form distinct patterns, with the exception of some moderately distinct very dark-brown transverse bands at the midpoint of thigh, tibia and ankle. Dorsal surface of hands and feet pale buff with extensive pale-brown flecking. Venter pale buff with fine tan maculations coalescing on the throat, becoming scattered posteriorly and less dense around the vent and thighs, giving the throat an overall darker appearance than the belly and thighs; ventral surfaces of limbs buff, almost translucent, with scattered pale-tan flecks, palmar surfaces unpigmented; plantar surfaces largely unpigmented with light-tan speckling.
Variation. Summary measurements for the type series are given in Table 1 View TABLE 1 . The paratypes are of similar dimensions to the holotype, although all are slightly larger (SUL 13.7–14.7 mm vs. 13.5 mm). All specimens have a dorsal colouration including a longitudinally oriented dark-brown hourglass and paler-brown triangular lateral markings, and a pale-buff venter with extensive light-brown speckling. The paratypes have slightly paler dorsal colouration than the holotype (particularly SAMA R67519 and SAMA R67517); most also differ in having a distinct thin grey dorsal stripe extending from snout tip to urostyle tip (except SAMA R67518). The tympanic annulus is indistinct in all specimens, although a faint anterio-ventral ridge is sometimes apparent. A distinct palebuff medial stripe extends the full length of the venter on paratypes SAMA R67517 and SAMA R67514. The slope of the loreal region varies slightly among specimens, from slightly concave to nearly flat. Tuberculation in all paratypes is minimal and generally restricted to the flanks, a few low tubercles antero-lateral to the eyes are apparent in some specimens.
The juvenile specimen (SAMA R67515) has the same dorsal and lateral colour pattern as adults, but the venter is much paler and almost entirely lacks flecks of light-brown pigmentation. The snout of the juvenile is also proportionally shorter than those of mature specimens, (SL/SUL 0.20 vs. an average of 0.23) while the eye diameter is proportionally larger (ED/SUL 0.13). Tibia length, head width and internarial distance are also relatively larger in the juvenile specimen. These data suggest that the elongated snout of this species undergoes positive allometric growth.
Appearance in life. Photographs in life ( Figure 2 View FIGURE 2 ) of three specimens (SAMA R67516, R67519, R67520) taken at night, show a greater degree of contrast between light and dark brown dorsal and ventral areas than shown in preservative. The dorsal hourglass marking shows greater tonal variation than in preservative, sometimes including reddish-brown patches within the darker areas. Venter tinged grey on the throat grading to pink on the belly, with extensive fine reddish-brown maculations and scattered large pale grey-blue flecks ( Figure 3 View FIGURE 3 ), grey medial stripe more clearly visible than in preservative. Iris black with extensive small bronze flecks, pupil horizontal, encircled by a thin reddish-orange ring. Dorsal surfaces of torso and legs with numerous small tubercles of varying size that are less apparent in preservative.
Advertisement call. Three full call sequences produced by SAMA R67520 at an air temperature of 25.5o C were recorded on 29 March 2008 ....... Calls are groups of 3–5 rather scratchy notes, sounding to the ear like: ‘crik..crik..crik..crik............crik..crik..crik..crik..’, repeated regularly, with each call separated from the next by 1.6– 6.2 s (mean = 2.2; SD = 0.72; n = 50). However only three of 50 inter-call intervals are more than 2.5 s long, and with these outliers removed the range is reduced to 1.6– 2.5 s (mean = 2.1; SD = 0.18; n = 47). The three call sequences last 36.5– 55.8 s (n = 3) and contain 14–22 calls, each lasting 0.3– 0.5 s (mean = 0.40; SD = 0.077, n = 53) and produced at a rate of 0.36–0.38/s (mean = 0.37; SD = 0.01). Each call contains 3–5 (mean = 4.2; SD = 0.6; n = 53) distinctly pulsed notes. Note repetition rate within calls is 7.2–13.3/s (mean = 8.9; SD = 0.8; n = 41) but only one of the 41 calls of sufficient quality for analysis has a note repetition rate of more than 10/s. Individual notes are 0.014– 0.042 s long (mean = 0.028; SD = 0.005; n = 107) and contain 4–9 (mean = 5.6; SD = 1.0; n = 68) pulses. Pulse repetition rate within notes is 133–319 pulses/s (mean = 183.6; SD = 47.3, n = 68). Dominant frequency of the three calls is 4213–4519 Hz (mean = 4370; SD = 152.8; n = 3). A series of calls is illustrated in Figure 4.
Etymology. The specific name gracilirostris is a feminine compound adjective, taken from the Latin “ gracilis ” meaning slender, in its combining form “ gracili– ”, and Latin “ rostrum ” meaning beak or snout, which in its adjectival combining form is “ –rostris ”.
Natural history and distribution. C. gracilirostris is known from four locations in primary foothill and lower-montane rainforest on the southern side of the Central Cordillera at altitudes ranging from 213–1368 m; this is the widest altitudinal range known for a Choerophryne species. Males were primarily found calling during rain at night, from within leaf litter or rotting logs within 20 cm of the ground. No other Choerophryne species were found in sympatry with the new species; however Choerophryne burtoni is known from higher altitudes (1600 m a.s.l.) in the Muller Range ( Kraus and Allison 2009b; Richards and Dahl 2009).
Molecular differentiation. A DNA sequence of a fragment of the mitochondrial 16S rRNA gene of the new species, amplified using the primers 16SL3 and 16SAH from Vences et al. (2003) has been made available on Genbank (accession number KF561136 View Materials ). Comparisons to homologous data for C. amomani , C. arndatorum , C. microps and C. nigrescens (from Köhler and Günther 2008) using the Species Delimitation plugin for Geneious Pro 5.5.2 software revealed uncorrected sequence divergence from all taxa to be between 16.5–21.0%. These molecular data support the evolutionary distinctiveness of C. gracilirostris sp. nov from all other taxa, including the morphologically similar C. arndatorum (e.g. Vieites et. al. (2009) suggested a guideline of>3% uncorrected pairwise genetic divergence in 16S rRNA for identification of candidate species).
FIGURE 4. Advertisement call of Choerophryne gracilirostris sp. nov. (SAMA R67520) recorded at a temperature of 25.5o C. A) Five consecutive calls from a longer sequence, showing waveform (top), spectrogram (bottom) and frequency distribution (left); B) Waveform (top) and spectrogram (bottom) of the third (middle) call illustrated in A, showing distinctly pulsed nature of the five notes.


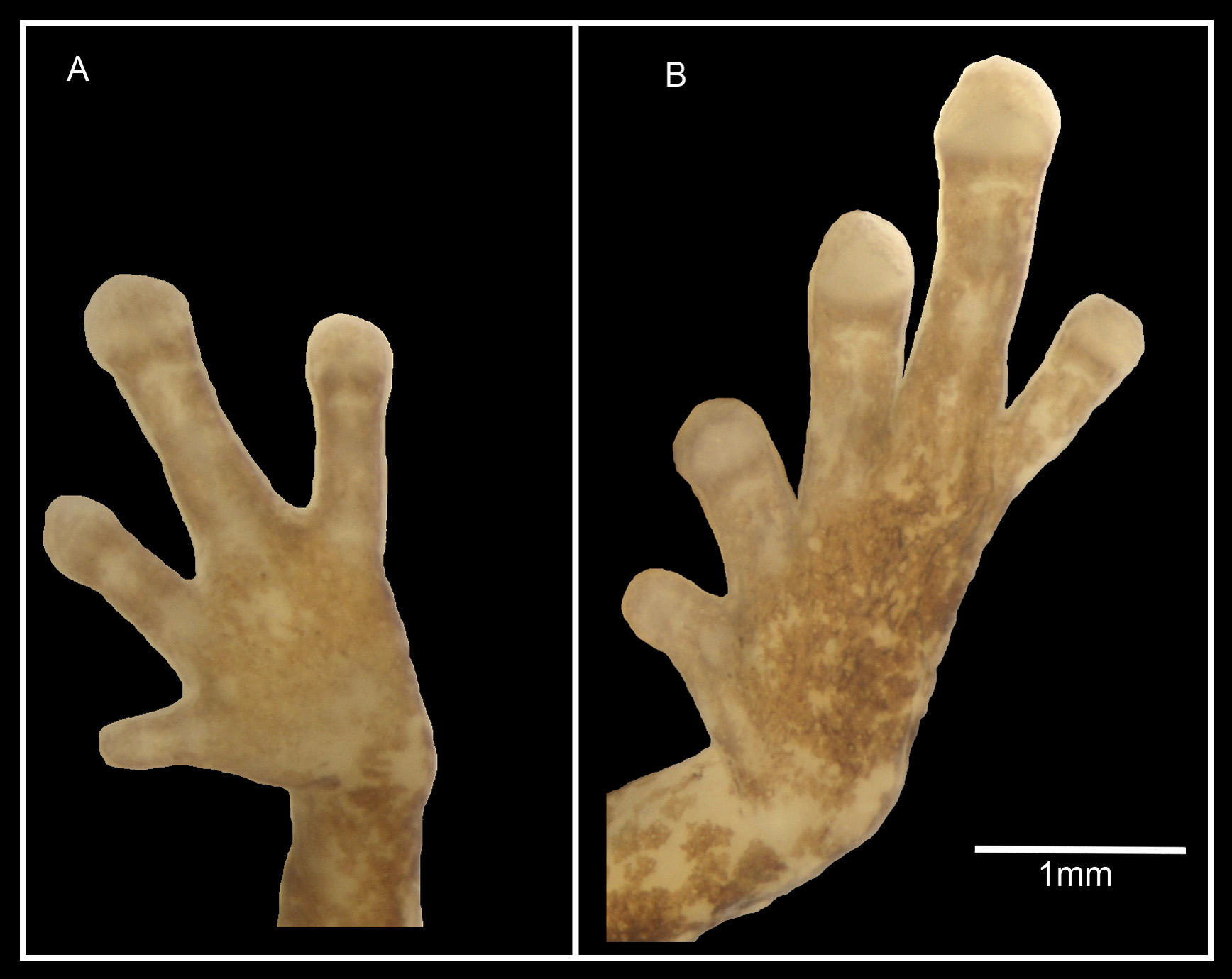

TABLE 1. Summary of measurements (mm) for Choerophryne gracilirostris sp. nov.
| Reg# | HOLOTYPE R67520 | PARATYPE R64986 | PARATYPE R67516 | PARATYPE R67517 | PARATYPE R67519 | PARATYPE R67514 | PARATYPE R67518 | R67515 |
|---|---|---|---|---|---|---|---|---|
| Sex | Male | Male | Male | Male | Male | Male | Male | Juvenile |
| SUL | 13.5 | 14.0 | 14.4 | 13.7 | 13.7 | 14.7 | 13.9 | 8.2 |
| TL | 5.4 | 5.5 | 5.3 | 5.4 | 5.2 | 5.6 | 5.4 | 3.3 |
| F1D | 0.2 | 0.2 | 0.2 | 0.3 | 0.3 | 0.2 | 0.3 | 0.1 |
| F3D | 0.5 | 0.5 | 0.4 | 0.4 | 0.5 | 0.5 | 0.5 | 0.2 |
| T1D | 0.4 | 0.4 | 0.4 | 0.3 | 0.4 | 0.4 | 0.4 | 0.1 |
| T4D | 0.6 | 0.5 | 0.6 | 0.8 | 0.7 | 0.8 | 0.8 | 0.4 |
| HL | 4.6 | 5.0 | 4.9 | 5.1 | 4.8 | 5.3 | 4.9 | 2.9 |
| HW | 4.7 | 5.1 | 5.3 | 5.0 | 4.6 | 5.3 | 4.8 | 3.1 |
| SL | 3.1 | 3.6 | 3.2 | 3.2 | 3.0 | 3.1 | 3.1 | 1.7 |
| ED | 1.3 | 1.4 | 1.5 | 1.5 | 1.5 | 1.5 | 1.6 | 1.1 |
| OHG | 1.2 | 1.1 | 1.1 | 1.0 | 0.9 | 0.9 | 1.0 | 0.5 |
| END | 2.1 | 3.2 | 2.2 | 2.0 | 1.9 | 2.2 | 1.9 | 1.1 |
| IND | 0.8 | 0.8 | 0.8 | 0.8 | 0.8 | 0.8 | 0.8 | 0.6 |
| SAMA |
South Australia Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |