
Agalychnis terranova, Rivera-Correa, Mauricio & Duarte-Cubides, Felipe, 2013
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3636.1.4 |
|
publication LSID |
lsid:zoobank.org:pub:D60DF231-A618-4AA6-808D-834AEA3EE8F0 |
|
DOI |
https://doi.org/10.5281/zenodo.5689279 |
|
persistent identifier |
https://treatment.plazi.org/id/0386327E-FFE8-FFA6-A7D2-8798ADA0FE4C |
|
treatment provided by |
Plazi |
|
scientific name |
Agalychnis terranova |
| status |
sp. nov. |
Agalychnis terranova sp. nov
( Figs. 3 – 5 View FIGURE 3 View FIGURE 4 View FIGURE 5 )
Holotype. MHUA-A 7316, adult male, Colombia, Departamento de Antioquia, Municipio de Sonsón, Corregimiento La Danta, Vereda La Danta, 05°50’8.04’’N, 74°50’16.55’’W, 380 m. a.s.l., collected on April 15, 2011, by Felipe Duarte.
Paratypes. MHUA-A 7317 – 7319, three adult males and MHUA-A 7320 subadult, collected with the holotype. ICN 42966 – 42969, four adult males, Colombia, Departamento de Cundinamarca, Municipio de Yacopí, Vereda Guadualito, 05º37’52’’N, 74º8’23’’W, 820 m. a.s.l., collected on November 1, 1995, by Jose Vicente Rueda, Juan Manuel Renjifo and Freddy Castellanos. ICN 42970 – 42971, two adult males, Colombia, Departamento de Cundinamarca, Municipio de Yacopí, Vereda Cabo Verde, collected on October 29, 1995, by Jose Vicente Rueda, Juan Manuel Renjifo and Freddy Castellanos. MHUA-L 0 198, 33 tadpoles, Colombia, Departamento de Antioquia, Municipio de Sonsón, Corregimiento La Danta, Vereda La Danta, 05°50’8.04’’N, 74°50’16.55’’W, 380 m. a.s.l., collected during April 15 – 18, 2011, by Felipe Duarte. MHUA-L 0 199, two tadpoles, Colombia, Departamento de Antioquia, Municipio de Sonsón, Corregimiento La Danta, vereda La Danta, 05°50’8.04’’N, 74°50’16.55’’W, 380 m. a.s.l., collected on February 15, 2012, by Eliana Muñoz.
Diagnosis. We assigned the new species to the genus Agalychnis and the A. callidryas group on the basis of our phylogenetic results ( Fig. 1 View FIGURE 1 ). Our hypothesis is morphologically supported by the presence of the two putative synapomorphies in the new species: iris red and palpebral membrane reticulated with gold (Faivovich et al. 2010). The new species is characterized and defined by the following combination of characters ( Figs. 2 – 3 View FIGURE 2 View FIGURE 3 ): (1) medium size for the genus (SVL 42.5 to 50.8 mm in adult males, 67.4 in one adult female); (2) slender body, head wider than body; (3) snout rounded in dorsal view and slightly sloping in lateral view; (4) loreal region slightly concave; (5) iris in life red wine with black reticulations; (6) pupil vertical, palpebral membrane with golden reticulations; (7) discs relatively wide, webbing formulae of fingers I (2 1/2–2 2/3) – 2 1/ 2 II (2–2+) – (2 2/3– 3) III (3–3-) – (2 1/2–2 2/3) IV), (8) webbing formulae of toes I (2–2+) – (2 1/3–2 1/2) II (2–2+) – (3+–3 1/3) III (2-–2+) – (3–3+) IV (3 2/3–3+) – (2-–2) V; (9) dorsally green with or without white warts; (10) concealed surfaces of arm, forearm, thigh, tibia, tarsus, and digits light orange; (11) flanks light orange or orange with small white warts (12) ventral surfaces creamy in life (13) males with nuptial pad covering medial margin of prepollex with a distal expansion that partially covers the dorsal surface of the metacarpus and with dark brown colored epidermal projections; (14) dorsal skin slightly coarse; (15) parotoid glands absent; (16) tympanum rounded with tympanic annulus distinct and upper edge covered by a supratympanic ridge; (17) inner ulnar and tarsal fold white present, both with smooth margin; (18) heel tubercles and calcars absent; (19) cloacal flap long, cloacal opening directed posteroventrally at level of ventral surfaces of thighs; cloacal sheath very short.
Comparison with other species. Agalychnis terranova is readily distinguished from the other species of A. callidryas group by the orange flanks with small white warts. Additionally, A. terranova differs from A. annae by the presence of red iris, absence of light blue hue in the flanks (iris orange and light blue hue present in A. annae ); from A. callidryas by absence of yellow vertical bars on the flanks on background blue or brown (present in A. callidryas ); from A. moreletii and A. spurrelli by having less extensive webbing that does not reach beyond the distal subarticular tubercles between fingers III and IV and toes III and IV (webbing beyond distal subarticular tubercles in A. moreletii and A. spurrelli ); from A. saltator in its larger size, without dark transverse stripes on the dorsum and, hidden areas dark blue or purple absent (presents in A. saltator ). Furthermore, A. terranova is readily distinguished from A. aspera , A. buckleyi , A. granulosa , A. hulli , A. lemur , A. medinae , and A. psilopygion by their pale silvery bronze or cream iris (red iris in A. terranova ), webbing on hands and feet absent (not extensive but present in A. terranova ), and dorsal skin very thick and densely granular (slightly coarse in A. terranova ). Furthermore none of these latter species has gold reticulated membrane palpebral (present in A. terranova and other A. callidryas group members); from A. dacnicolor by having golden iris with black thick reticulations (red iris in A. terranova ) and membrane palpebral not reticulated (gold reticulated in A. terranova ). Agalychnis danieli has iris golden, irregular black spots in lips, thighs, inner and outer margin of the arms and legs, and a calcar tubercle developed present (red iris present, blotches and calcar tubercle absent in A. terranova ).
Description of the holotype. Body slender; head depressed, slightly wider than longer, head width 37% SVL; head length 32% SVL; snout rounded in dorsal view and sloping in lateral view from eyes to nostrils, then further inclined to tip of snout to the top of the head; lips flared; nostrils placed laterally, slightly protuberant and situated at about three-fourths the distance from the eyes to the tip of the snout; internarial area depressed; canthus rostralis broadly rounded, barely distinct; loreal region slightly concave; eye large, protuberant, directed anterolaterally, eye diameter approximately 34% of head length; internarial distance 20% of head width and 69% of eye diameter. Tympanum and tympanic annulus visible, round, laterally positioned, distinct dorsally, barely hidden by a thin dermal fold which extends in a curve from upper posterior corner of tympanum until the insertion of arm; tympanum diameter about one-half of eye length, nearly one-third of distance eye to nostril and separated from eye by distance equal to one-half of its diameter. Tongue lanceolated, free on the posterior half; choanae large, oval; dentigerous processes of vomers oblique and posteromeadially directed, between choanae, with processes separated by a relative distance of 40% of the diameter of choanae, each bearing six teeth. The vocal slits lie along the inner posterior margin of the jaw. The vocal sac is single, median, subgular, and not noticeably distensible.
Upper arm long and slender; forearms long and robust, with thin and smooth dermal ridges on ventrolateral edge of forearm from elbow to disc of fourth finger; fingers short, with circular and well-developed terminal discs, not opposable; disc diameter on the third finger equal to that of tympanum; finger lengths I <II <IV ≤ III ( Fig. 5 View FIGURE 5 C); subarticular tubercle large size, ovoid, conical; supernumerary tubercles present, rounded, sub-conical, few and non-distinctive; inner metacarpal tubercle large, simple, elliptical, at the base of finger I and covering the metacarpal; outer metacarpal tubercle absent, but a tubercle present located almost in the center of the palm, round, simple, flat; nuptial pad covering medial margin of prepollex with a distal expansion that partially covers the dorsal surface of the metacarpus and with dark brown colored epidermal projections. Webbing formula of finger I 2 1/2 –2 1/ 2 II 2–2 1/ 2 III 3-–2 1/ 2 IV
Legs long and slender; thigh and tibia of the same length; the smooth tarsal fold from heel to disc on the fifth toe; foot slender with toe lengths I <II <III <V <IV ( Fig. 5 View FIGURE 5 D), with well-developed discs, circular, slightly smaller than those of fingers discs; toes not opposable; inner metatarsal tubercle medium sized, simple, flat, ovoid; outer metatarsal tubercle absent; subarticular tubercles medium sized, ovoid, and prominent; supernumerary tubercles are present on the proximal segments of all but the first toe; webbing formulae of toes I 2 +–2 1/ 3 II 2–3 1/ 3 III 2+– 3 IV 3+– 2 V.
Dorsal skin smooth with two medium rounded warts; granular flanks with small scattered warts; gular region smooth; belly and proximal area of thighs areolate; ventral surfaces of arms and limbs, smooth; cloacal region granulated. Cloacal flap long; cloacal opening directed posteroventrally at level of ventral surfaces of thighs. Paratoids and dorsolateral glands absent.
Color in life of holotype. Dorsal surfaces of body, forearms, legs, loreal, and tympanic regions green; orangegreen color covering most of outer finger and outer toe, others fingers and toes and upper arms light orange; light orange dermal ridges on forearms and dermal flap on heel white; concealed surfaces of arm and thigh light orange; flanks light orange with white warts; hands, feet and ventral surfaces of limbs orange; throat and cloacal area are white and the belly is creamy white; nuptial excrescence dark brown; vertical pupil black, iris red with black reticulations, and golden reticulations extending widely throughout palpebral membrane. Color in life of a paratype shown in Fig. 3 View FIGURE 3 .
Color in preservative of holotype. Dorsal surfaces of body, forearms, legs, loreal, and tympanic regions sky blue, dorsum with two white warts of different sizes; upper arms, fingers, and toes cream; iris light cupreous; flanks, inguinal region, internal and external sides of tibia, and internal side of tarsus cream; ventral surfaces cream; nuptial excrescence dark brown ( Fig. 2 View FIGURE 2 ).
Measurements of the holotype (in mm). SVL 47.9; HL 15.2; HW 17.3; ED 5.2; END 4.6; NSD 2.3; IND 3.0; AMD 12.2; TD 3.9; IOD 13.3; THL 27.1; TBL 26.1; TAL 15.5; FL 17.8; TFD 2.8; FFD 2.3
Variation. Measurements of the type series are summarized in Table 2 View TABLE 2 . No sexual variation between the morphology of males and females, except for the sexually dimorphic structures (i.e. nuptial pads and vocal slits in males) and females are about 25% larger than males in SVL. Dorsal skin slightly coarse with scattered white warts without dark ring in some individuals: one wart (ICN 54248, MHUA-A 7318–7319), three warts (MHUA-A 7317) four warts (ICN 42967, 42970, 54249). Number of vomerine teeth range from 3 to 8, sometimes asymmetric (left/ right): 4/3 (MHUA-A 7320), 4/4 (MHUA-A 7319), 4/5 (ICN 42970), 4/7 (MHUA-A 7318), 5/5 (ICN 42967, MHUA-A 7317), 6/7 (ICN 54248), 7/8 (ICN 54252). The white warts of flanks and ventral thighs are less numerous and more spaced in larger specimens, and some warts have minute melanophores on the margin. Nuptial excrescences less developed in ICN 54249 and undetectable in ICN 42970 and MHUA-A 7320. Variation in webbing formulae of fingers I (2 1/2–2 2/3) – 2 1/ 2 II (2–2+) – (2 2/3– 3) III (3–3) – (2 1/2) IV) and variation of webbing formulae of toes I (2–2+) – (2 1/3–2 1/2) II (2–2+) – (3+–3 1/3) III (2-–2+) – (3–3+) IV (3 2/3–3+) – (2-–2) V. In preservative, some individuals possess darker blotches, almost deep purple on different areas of dorsum and legs.
Tadpole description. Morphometric measurements are shown Table 3; external morphology in life ( Fig. 5 View FIGURE 5 ). Body ovoid in lateral view, elliptic in dorsal view. Body length slightly less than half total length, body two times longer than wide, 2.3 times longer than tall and 1.1 times wider than tall. Snout shape rounded to truncated in dorsal profile and rounded in lateral profile. Nostril dorsolateral, directed anteriorly, elliptical with slightly flattened dorsal edge, margin in nostrils with rim; closer to tip of snout than eyes. Eyes dorsolateral and directed laterally; relatively large, eye diameter 40% of the maximum height of the body. Spiracle ventral sinistral to the midline, centripetal with wall absent and an opening 30% of the width of the oral disc. Neuromasts of the lateral line system not evident. Vent tube short, dextral, opening postero-laterally directed, connected along all its length with the ventral fin on the ventral edge. Caudal musculature robust anteriorly and gradually thinning towards the tip; dorsal fin emerging posteriorly to the body tail-junction, not extending onto body, more tall in the two thirds of the total length of the tail; ventral fin is slightly deeper than the dorsal fin and has about equal depth on the middle third length of tail, emerging anteriorly to vent tube and narrowing posteriorly. Hind limb length (stage 39) equals 32.4% of total length, many melanophores outer in femur and radio-ulna present; plantar tubercle is more conspicuous than subarticular tubercles and dorsum of phalanges are brownish; vestigial webs are in fingers III–IV, IV–V.
Oral disc medium, width 50% of maximum body width, anteroventral, directed anteroventrally, not emarginated; the median part of anterior labium is bare, width 50% of oral disc width; median part of posterior labium is bordered by single row of papillae; the rest of the mouth is bordered by two to four rows of papillae, laterally abundant submarginal papillae ( Fig. 5 View FIGURE 5 A). Upper jaw moderately robust, keratinized, forming a broad arch with slender and large lateral process, expanded distally. The lower jaw is more slender and V-shaped. Both jaws have fine serrations, wide at the base; jaw sheath serrations closely spaced. Two rows of labial teeth on the anterior labium, three rows of labial teeth on the posterior labium; labial tooth row formula (LTRF) 2(2)/3(1), longitude of gap in A2 about 6% of total length of the row, gap in P1 narrowest, inconspicuous; rows of labial teeth on anterior labium equal in length and extend nearly to lateral papillae; second row of the labial teeth on posterior labium (P2) 6.5% shorter than first row (P1); third row of the labial teeth on posterior labium (P3) 4% shorter the second row (P2).
Color in life of all tadpoles. Background color of dorsum and flanks ocher-brown with blotches and olive brown and silver melanophores present. Oral disc with many spaced melanophores. Parietal peritoneum iridescent platinum-yellow. Caudal musculature light brown with blotches olive brown. Dorsal and ventral tail fins with transparent background highly vascularized with scattered red spots and scattered dark pigmentations that persist in preserved specimens with dendritic appearance. Inner ridges of ventral and dorsal fin above tail muscle with dense melanophores present in the proximal area of tail length. Iris gold.
Ontogenic changes. At stages 25 and 26 the body form is more depressed that at stage 27. The cement gland is not observed in the early stages. Melanophores at stages 25 to 28 are uniformly distributed, after stage 29 numerous blotches in the flanks and caudal musculature present. At stage 25 the papillae are widely spaced, others stages have numerous papillae with reduced space. Early stages (25 to 31) have diminutive and conspicuous spots above vental tube. The tail-muscle junction is more protuberant after stage 31, and the wall of the vental tube is largest after stage 33. In later stages (34 to 39) numerous melanophores appear closer to the tail-muscle junction. Finally, at stages 38 and 39 length of gap in anterior labium increases. The LTRF remains unchanged throughout development.
Etymology. The specific epithet terranova comes from the Latin words terra (Earth) and novus (new), meaning that which comes from a new land. Here we attribute the name, referring to the distribution of the new species in the Magdalena River valley of Colombia, previously not known to host any member of the genus Agalychnis .
Geographic distribution and natural history. Agalychnis terranova has been recorded at four localities in the Andean foothills between 380 and 900 meters along the Middle Magdalena River valley ( Fig. 6 View FIGURE 6 ). The region is characterized by a rugged relief covered by tropical humid forests (bh-T) sensu Holdridge (1987). Type locality is mostly represented by forest remnants having a plant community dominated by plant species such as Cecropia insignis, Clusia palida, Cupania americana, Gleoespermum longifolium, Morisona oblongifolia, Inga umbellifera, Talisia oliviformis, Therocarpus officinali and Trichilia moschata. The specimens from the type locality were observed active at night, calling from vegetation between 2 to 3 meters high, near artificial ponds. Tadpoles were collected in an artificial pool of 4 x 5 m length, forming schools or remaining motionless at an oblique angle to the sheet of water. The artificial pool where the tadpoles were observed was dominated by plants of the genus Musa . Phyllomedusa venusta tadpoles were also observed at the site. At the time of collection we did not observe photosensitive reaction in the tails of larvae and also did not register clutches or adults in amplexus. Other amphibians found in sympatry were Craugastor raniformis , Dendrobates truncatus , Engystomops pustulosus , Hypsiboas boans , Leptodactylus bolivianus , Phyllomedusa venusta , Pristimantis taeniatus, Rhinella gr. margaritifera, Sachatamia punctulata , Scinax rostratus and Smilisca phaeota . A KOH test revealed positive presence of pterorhodin in the skin of Agalychnis terranova .
Remarks. Recently Faivovich et al. (2010) assessed the phylogenetic relationships of the subfamily Phyllomedusinae based on a comprehensive molecular analysis, including 75% of the known species. In their taxonomic proposal, Hylomantis and Pachymedusa were synonymized with Agalychnis . In addition, Faivovich et al. (2010) proposed the Agalychnis callidryas species group, a well-supported clade including all of the species traditionally assigned to the genus Agalychnis (sensu Duellman 1970; Faivovich et al. 2005). Morphologically, the A. callidryas group is characterized by a gold reticulated palpebral membrane ( Fig. 7 View FIGURE 7 ) and a red hue of the iris, and both characters have been suggested as putative synapomorphies (red hue with a subsequent transformation into orange iris in A. annae ). Furthermore, the study by Faivovich et al. (2010) suggested that the webbing on the hands and feet, which is more extensively developed than in other congeners outside of the group, could be an additional putative synapomorphy. These character states also are present in A. terranova , thus supporting the hypothesis of such character as a putative synapomorphy for the Agalychnis callidryas group, given that membership of A. terranova in the group is confirmed by independent (molecular) data.
In addition to the morphological evidence, the validity of A. terranova as a distinct species is also supported by genetic divergence given that the uncorrected genetic distance between this taxon and its sister species A. callidryas is higher than between other congeneric sister species, for example A. annae and A. moreletii (Table 1).
The phylogenetic position of A. terranova represents an opportunity to start building hypotheses about Agalychnis biogeography. Among the A. callidryas group, only A. spurrelli and A. callidryas reach the northwestern region of South America and are restricted mostly to the lowland forests of Choco. The records of two individuals of A. callidryas in an isolated population in the Botanical Garden of Cartagena, Department of Bolívar, northern Colombia (Ruiz-Carranza et al. 1997) could represent an introduced population established by accident via the ornamental plant trade. To date, Agalychnis terranova is the only species of this genus in the Magdalena River valley. The lowland rainforest in the Pacific coast of Colombia and Ecuador, the northern of the Cordillera Occidental and Central in Colombia and the humid portion of the Magdalena river valley has been considered a single biogeographic province (Hernandez-Camacho et al. 1992). However, recent studies suggest that a potential biogeographic break might occur between the Chocó and the Magdalena lowland forests where apparently no geographic barriers can be distinguished (Daza et al. 2009, Saldarriaga-Córdoba et al. 2009, Pinto- Sánchez et al. 2011). This break will explain the presence of A. terranova in the Magdalena river valley and its sister taxa in the Choco region. Additional phylogeographic studies with a thorough sampling in northern Antioquia and the Chocó region will test this idea and contribute to understand the historical processes leading to the biotic divergence between Middle America lowlands and the Magdalena River valley regions.
Despite that the Magdalena River valley is relatively well-studied, A. terranova is known from only four localities and biological collections remain under-represented. This could be because the typical natural habitat of this species is the forest canopy, like some other species of the genus (Roberts 1994; Wells 2007). This would indicate that random encounters are associated with reproductive activity near the ground, because this species requires lotic water bodies for the development of the larval stages. Future studies on the reproductive biology will be needed to understand aspects of the natural history of this species.
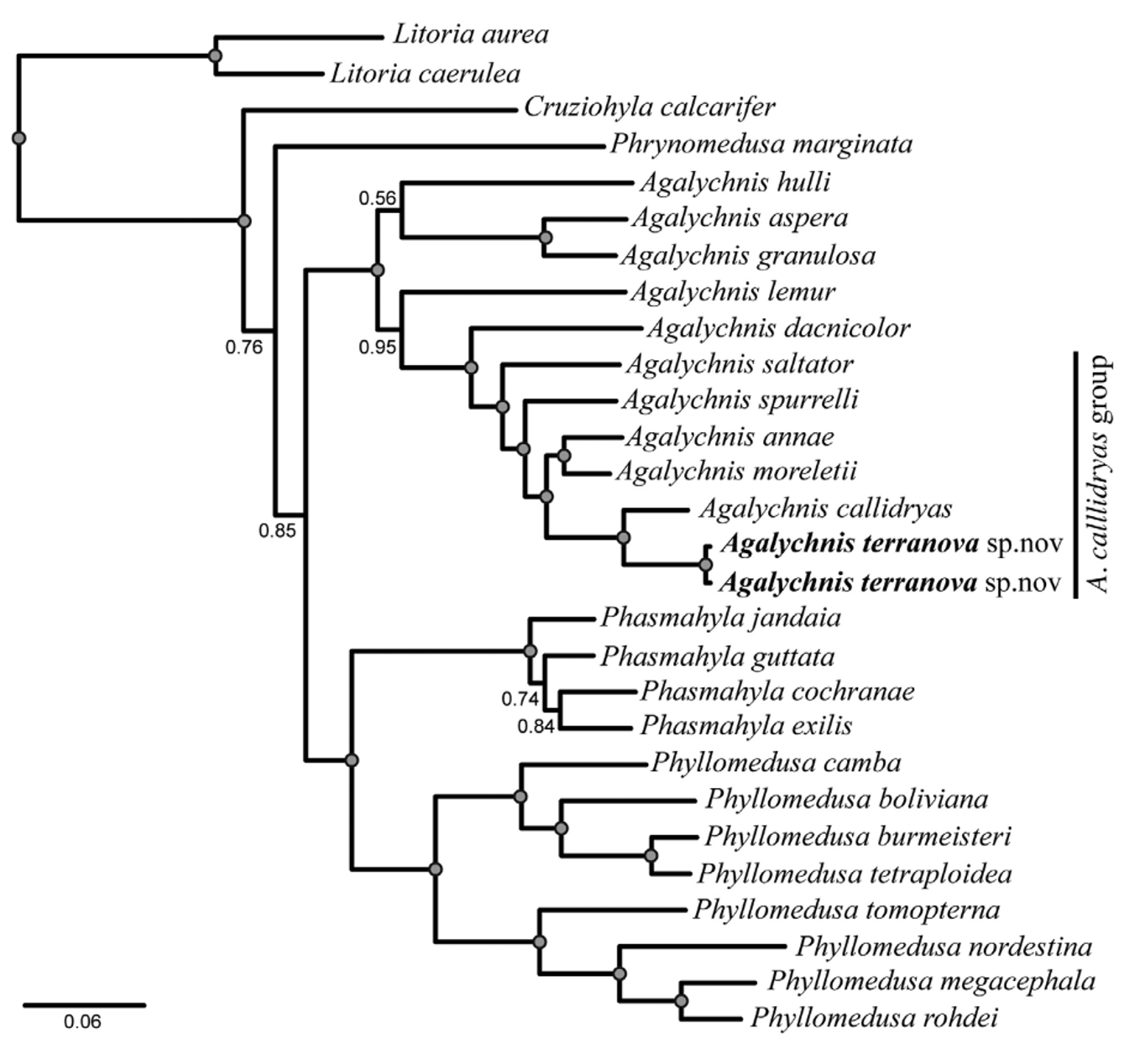






TABLE 2. Measurements (in mm) of adults Agalychnis terranova sp. nov. See text for abbreviations; min, minimum value; max, maximum value; mean, arithmetic mean; SD, standard deviation.
| Males (n=14) | Female (n=1) | ||
|---|---|---|---|
| Measurement | min max mean | SD | |
| SVL | 42.5 50.8 46.9 | 2.63 | 67.4 |
| HL | 13.9 18.9 16.1 | 1.63 | 20.1 |
| HW | 13.4 17.6 15.1 | 1.52 | 22.8 |
| ED | 4.6 5.9 5.4 | 0.38 | 6.0 |
| END | 4.2 5.1 4.3 | 0.30 | 5.9 |
| NSD | 1.8 2.7 2.2 | 0.25 | 2.8 |
| IND | 3.3 4.1 3.8 | 0.22 | 4.6 |
| AMD | 9.4 11.7 10.4 | 0.65 | 13.1 |
| TD | 2.3 3.4 2.8 | 0.31 | 3.9 |
| FAL | 9.8 13.0 11.4 | 0.99 | 16.4 |
| FAB | 3.1 4.5 3.7 | 0.37 | 4.0 |
| HAL | 12.0 13.9 13.0 | 0.52 | 18.3 |
| THL | 22.6 27.1 24.9 | 1.36 | 37.6 |
| TBL | 22.8 26.1 24.6 | 0.82 | 36.4 |
| TAL | 13.1 15.8 14.6 | 0.71 | 21.4 |
| FL | 15.2 18.9 16.9 | 1.06 | 24.4 |
| TFD | 2.4 2.9 2.6 | 0.18 | 4.0 |
| FFD | 2.2 2.5 2.4 | 0.09 | 3.6 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.