
Physalaemus araxa, Leal & Zornosa-Torres & Augusto-Alves & Dena & Pezzuti & Leite & Lourenço & Garcia & Toledo, 2021
|
publication ID |
https://doi.org/10.5852/ejt.2021.764.1475 |
|
publication LSID |
lsid:zoobank.org:pub:AE566208-36CD-4FB6-A6C9-EDECB157B9B1 |
|
DOI |
https://doi.org/10.5281/zenodo.5559854 |
|
persistent identifier |
https://treatment.plazi.org/id/5F59A249-8820-4EAD-8BDA-9603A8F3B4EE |
|
taxon LSID |
lsid:zoobank.org:act:5F59A249-8820-4EAD-8BDA-9603A8F3B4EE |
|
treatment provided by |
Felipe |
|
scientific name |
Physalaemus araxa |
| status |
sp. nov. |
Physalaemus araxa sp. nov.
urn:lsid:zoobank.org:act:
Figs 1–4 View Fig View Fig View Fig View Fig , 5D View Fig , 6–7 View Fig View Fig ; Tables 1–6 View Table 1 View Table 2 View Table 3 View Table 4 View Table 5 View Table 6
Physalaemus sp. ( aff. maculiventris View in CoL ) – Zornosa-Torres et al. 2020: 4–6, 12–13.
Type material
Holotype BRAZIL • adult ♂; state of Espírito Santo, municipality of Ibitirama, Parque Nacional do Caparaó, Lagoa da Sombra ; 20.421778° S, 41.802500° W; 2505 m a.s.l.; datum WGS84; 5 Oct. 2017; C. Zornosa-Torres, G. Augusto-Alves and S. Dena leg.; ZUEC-AMP 24095 . ( Figs 1–2 View Fig View Fig ; Table 1 View Table 1 ) GoogleMaps
Paratypes BRAZIL • 6 adult ♂♂; same collection data as for the holotype; ZUEC-AMP 24094 , 24096 to 24100 GoogleMaps • 2 adult ♂♂; same locality as for the holotype; 26 Nov. 2014; F.S.F. Leite leg.; UFMG 17250 View Materials , 17251 View Materials GoogleMaps • 4 adult ♂♂; state of Minas Gerais, municipality of Alto Caparaó, Parque Nacional do Caparaó, Três Lagoas ; 20.430972° S, 41.802611° W; 2505 m a.s.l.; datum WGS84; 10 Oct. 2017; C. Zornosa-Torres, G. Augusto-Alves and S. Dena leg.; ZUEC-AMP 24117 to 24120 GoogleMaps .
Additional material examined
BRAZIL • 3 juveniles raised in captivity from wild-collected tadpoles; same locality as for the holotype; 25 Jan. 2018; C. Zornosa-Torres, G. Augusto Alves and C. Nunes de Almeida leg.; ZUEC-AMP 24403 to 24405 GoogleMaps • 1 lot of 25 tadpoles; state of Minas Gerais, municipality of Alto Caparaó, Parque Nacional do Caparaó, Três Lagoas ; 20.430972° S, 41.802611° W; 2505 m a.s.l.; datum WGS84; 6 Dec. 2017; C. Zornosa-Torres, G. Augusto Alves and R. Santos leg.; ZUEC-AMP 24214 GoogleMaps .
Diagnosis
Physalaemus araxa sp. nov. is distinguishable from all congeneric species by having the following combination of character states: (1) presence of an arrow-shaped blotch on the dorsum of the body ( Fig. 1A View Fig ); (2) tarsal tubercle absent ( Fig. 2D View Fig ); (3) gular region and chest predominantly yellow in live individuals, pale cream in preserved specimens ( Figs 1B View Fig , 3B–C View Fig ); (4) belly with dark vermiculation pattern on a pale cream, slightly bluish background in live individuals ( Figs 1B View Fig , 3C View Fig ); (5) lack of aposematic coloration on ventral surface of hand and foot in live individuals ( Fig. 3C View Fig ); (6) intermediate size within the P. signifer clade (adult male SVL = 17.4–21.5 mm, Table 1 View Table 1 ); (7) presence of brown, divided, nuptial pad in males ( Figs 1A View Fig , 2C View Fig ); (8) supernumerary tubercles on foot absent ( Fig. 2D View Fig ); (9) tarsal fold absent ( Fig. 2D View Fig ); (10) texture of posterior region of belly and ventral surface of thigh smooth ( Fig. 1B View Fig ); (11) advertisement call duration from 69–304 ms ( Table 2 View Table 2 ); (12) tadpole with a proportionally large body BL/TL = 0.39–0.43 ( Table 4 View Table 4 ); (13) dextral vent tube ( Fig. 6G View Fig ); (14) dorsal and ventral fins of the same height ( Fig. 6A, H View Fig ; Table 4 View Table 4 ); (15) tail tip broadly rounded ( Fig. 6A, H View Fig ); (16) presence of submarginal papillae arranged in small rows; (17) A2 tooth row conspicuously longer than A1 ( Fig. 6D View Fig ); (18) absence of a dermal fold at the body-tail junction; (19) external margins of fins slightly convex ( Fig. 6A, H View Fig ); (20) gular region of tadpole convex.
Comparison with other species
Physalaemus araxa sp. nov. may be set apart from all the species of the Physalaemus cuvieri clade (sensu Lourenço et al. 2015) by simultaneously having (1) the presence of an arrow-shaped blotch on the dorsum of the body and (2) absence of tarsal tubercle ( Leal et al. 2020). (3) The gular region and chest predominantly yellow-colored in live individuals, pale cream in preserved specimens, distinguish the new species from all the other species of the P. signifer clade (gular region and chest predominantly dark brown in live and preserved individuals of those species). Additionally, live individuals of Physalaemus araxa sp. nov. have (4) belly with dark vermiculation on a pale cream, slightly bluish background, distinguishing it from P. angrensis , P. atlanticus , P. nanus , and P. spiniger (belly with flashy orange blotches in those species), from P. maculiventris (which has a pale chest and belly, with posterior region of belly and ventral surface of thigh showing bold black blotches), and from P. obtectus (which has belly with flashy red blotches). (5) The lack of aposematic coloration on ventral surface of hand and foot in live individuals distinguishes the new species from P. angrensis , P. atlanticus , P. spiniger (orange aposematic coloration present on ventral surface of hand and foot in live individuals of those species) and from P. claptoni , P. deimaticus , P. erythros (red aposematic coloration present on ventral surface of hand and foot in live individuals of those species). (6) By having adult males of intermediate size within the P. signifer clade (SVL = 17.4–21.5 mm), P. araxa sp. nov. is set apart from P. bokermanni (which is smaller, SVL = 15.3–17.0 mm) and from P. caete , P. camacan , P. moreirae , P. nattereri , and P. obtectus (which are larger, combined SVL = 22.3–50.6 mm). (7) The presence of a brown, divided, nuptial pad in males distinguishes P. araxa sp. nov. from P. claptoni (which has a nuptial pad not divided) and from P. rupestris (which has a white cream nuptial pad). (8) The lack of supernumerary tubercles on foot distinguishes P. araxa sp. nov. from P. angrensis , P. caete , P. camacan , P. crombiei , P. irroratus , P. moreirae , P. signifer , and P. spiniger (supernumerary tubercles present on the foot in those species). (9) The lack of a tarsal fold distinguishes P. araxa sp. nov. from P. atlanticus , P. bokermanni , P. camacan , P. crombiei , P. irroratus , P. nanus , P. obtectus , P. signifier , and P. spiniger (tarsal fold present in those species). (10) Texture of posterior region of belly and ventral surface of thigh smooth in P. araxa sp. nov. distinguishes it from P. camacan and P. irroratus (posterior region of belly and ventral surface of thigh granulated in those species). (11) The duration of the advertisement call of the new species ranges from 69–304 ms, setting it apart from P. angrensis , P. atlanticus , P. bokermanni , P. caete , P. camacan , P. claptoni P. crombiei , P. moreirae , P. rupestris , and P. signifer (which have longer advertisement calls, combined minimum advertisement call duration from 324–2,130 ms). (12) Tadpoles of P. araxa sp. nov. have a proportionally larger body (BL/TL = 0.39–0.43), differing from those of P. atlanticus , P. bokermanni , P. maculiventris , P. moreirae , and P. spiniger ( 0.34 in P. atlanticus , 0.35 in P. bokermanni , 0.33 in P. maculiventris , 0.27– 0.37 in P. moreirae , 0.37 in P. signifer , 0.34 in P. spiniger ). (13) Tadpoles with dextral vent tube distinguish the new species from P. atlanticus , P. caete , P. camacan , P. nanus , P. rupestris , and P. spiniger (vent tube medial in those species). (14) By presenting dorsal and ventral fins of the same height tadpoles of P. araxa sp. nov. differ from those of P. angrensis , P. atlanticus , P. caete , P. camacan , P. crombiei , P. erythros , P. irroratus , P. moreirae , P. nanus , P. rupestris , P. signifer , and P. spiniger (dorsal fin higher than ventral one in those species) and from P. caete (which has dorsal fin lower than ventral one). (15) The broadly rounded tail tip also differs tadpoles of P. araxa sp. nov. from most species of the P. signifer clade, such as P. atlanticus , P. bokermanni , P. caete , P. camacan , P. crombiei , P. maculiventris , P. spiniger (tail tip pointed in those species) and P. angrensis , P. erythros , P. irroratus , P. moreirae , P. nanus , P. rupestris , P. signifier (tail tip nearly rounded in those species). (16) Tadpoles presenting submarginal papillae arranged in small rows set P. araxa sp. nov. apart from the remaining species of the P. signifer clade (which have submarginal papillae scattered in the lateral portions of the oral disc), except from P. erythros which has a similar condition. (17) The A2 tooth row conspicuously longer than A1 distinguishes tadpoles of P. araxa sp. nov. from those of P. atlanticus , P. bokermanni , P. camacan , P. erythros , P. maculiventris , and P. moreirae (A1 = A 2 in those species) and from P. angrensis , P. caete , and P. spiniger (A1> A 2 in those species). Additionally, tadpoles of P. araxa sp. nov. also differ from those of P. maculiventris by (18) the absence of a dermal fold at the body-tail junction (dermal fold present in P. maculiventris ), by (19) the external margins of the fins slightly convex (fins markedly convex in P. maculiventris ) and by (20) the gular region convex (gular region straight in P. maculiventris ).
Etymology
The specific epithet ʻ araxa ʼ, is the combination of the Tupi-Guarani indigenous language words ʻaraʼ (meaning ʻworldʼ) and ʻeçaʼ (meaning ʻto seeʼ) meaning ʻthe first place where the sun can be seenʼ, in reference to the sunshine view on the top of hills ( Chiaradia 2008). Also used to indicate the highest mountain in a landscape. That is the specific case of the type locality of Physalaemus araxa sp. nov., found on the mountaintop of the highest hill of the Atlantic forest.
Description
Holotype ( ZUEC-AMP 24095 )
Adult male ( Figs 1–2 View Fig View Fig ), SVL 21.0 mm. Head slightly wider than long. Head width 38.0% SVL and length 36.0% SVL. Snout rounded in dorsal view and rounded to truncated in lateral view ( Fig. 2A– B View Fig ). Canthus rostralis distinct, rounded; loreal region slightly concave. Snout protruding beyond lower jaw. Nostril dorsolaterally oriented, faintly protruding. Internarial region flat; top of the head slightly concave. Eye slightly prominent, anterolaterally oriented, its diameter 5% larger than END. Tympanum indistinct externally. Supratympanic fold distinct, thick, extending from the posterior corner of the eye to the shoulder. Dentigerous process of vomer absent. Premaxillary and maxillary teeth absent. Choanae rounded, separated from each other by a distance as large as four times its diameter. Tongue elongated, constricted on its anterior third, wider on its posterior half, free around lateral and posterior margin. Vocal slit present, longitudinal, originating on the sides of the tongue in its anterior third, and extending towards the corner of the mouth. Vocal sac single and subgular, faintly differentiated externally. Dorsolateral fold present, weakly distinct, from the posterior corner of the eye to the inguinal region. Forearm hypertrophied in relation to upper arm; upper arm slender, short. Fingers thick, without webs, relative lengths I <II = III <IV; finger tips not expanded. Proximal subarticular tubercles large, simple, prominent, and rounded; distal subarticular tubercles present on fingers III and IV, approximately of the same size as the proximal ones; supernumerary tubercles large, rounded, low, more distinct in the left hand. Inner and outer metacarpal tubercles ovoid, large, prominent. Nuptial pad divided, densely covered by dark keratinized spicules, present on the dorsal and lateral surfaces of the thumb (except for the distal phalange) and on the internal surface of the internal metacarpal tubercle. Tibia length 39% SVL; foot length 60% SVL. Toes thick, without webs, relative lengths I <II = V <III <IV; toe tips not expanded. Subarticular tubercles distinct, simple, prominent, and rounded; supernumerary tubercles absent. Tarsal fold absent; tarsal tubercle absent. Inner metatarsal tubercle distinct, ovoid; outer metatarsal tubercle distinct, rounded. Inguinal gland well developed, oval. Cloacal opening directed posteriorly at upper level of thighs. Region below the cloaca with low and faintly distinct tubercles scattered, encroaching the thighs on its posteroventral edge, where tubercles become increasingly less distinct. In preservative, texture of ventral, lateral, hidden, and dorsal surfaces are smooth. Measurements of the holotype in Table 1 View Table 1 .
Color of the holotype preserved in alcohol 70%
In preservative ( Figs 1–2 View Fig View Fig ), dorsal background color of the body, head, and limbs dark brown. All dorsal blotches dark brown, darker than background color. Interorbital blotch triangular-shaped, one vertex over each eyelid, the third vertex connected with the tip of an arrow-shaped blotch medially located on the dorsum of the body. Middle of the arrow-head with a small light spot, posterior portion of the arrow extends transversely towards each inguinal gland. Urostyle region bears a longitudinal blotch with anterior margin poorly defined, centered by a longitudinal light blotch. Some poorly defined and irregularly shaped small blotches scattered through dorsal surfaces of head and body. Inguinal gland ca 90% covered by a black ocellus. Dorsal surfaces of thigh, tibia, and foot with transversal blotches. Many white dots scattered throughout dorsal surface of body and limbs, mainly outlining the more distinct blotches and the black ocelli over the inguinal gland. Heel, anterior part of knee, and forearm with a black blotch. Upper arm with a longitudinal black blotch covering its posterior surface and the elbow. Dorsolateral black blotches, dorsally outlined by small white dots, extending from the posterior margin of the eye through the supra tympanic fold and reaching the second third of the flank. Elongated black blotches extends from the tip off the snout to the eye, passing over the nostril and canthus rostralis. Irregularly shaped black blotches on the loreal region and upper lip. Region between eyes and the insertion of the arms, pale cream. Ventral surfaces of the limbs brown with scattered light spots; ventrolateral edge of forearm with a black blotch over glandular tissue. Black blotch above the cloaca, dorsally outlined by small white spots, curved down toward the back of the thighs. Gular region and chest predominantly pale cream, stained with very small dark dots. Ventral border of mandible brown colored, without distinct blotches. Belly with dark vermiculation pattern on a pale cream background. Iris dark brown with a faintly visible black vermiculation; pupil black and horizontal.
Color in life
Gular region, chest, axillary region, lower border of anterior half of the flanks, and region between eyes and the insertion of the arms yellow ( Fig. 3B–D View Fig ). Inguinal region pink posteriorly to the black ocelli and yellow around its anterior edge ( Fig. 3A, C–D View Fig ). Ventral surface of limbs pinkish brown ( Fig. 3C View Fig ). Background color of belly pale cream, slightly bluish ( Fig. 3C View Fig ). Overall dorsal background and blotches coloration varying from brownish to greenish, background color with some sparkled areas of cream and orange brown.
Variation
Measurements and proportions of 13 adult males are presented in Table 1 View Table 1 . In life, color of dorsal background of head, body, and limbs may vary from pale yellow to dark brown; some individuals had these areas, as well as the region between eyes and the insertion of the arms, stained by pinkish coloration in different levels of area size and color intensity. Yellow coloration present on gular region, chest, axillary region, lower border of anterior half of the flanks, and the region between eyes and the insertion of the arms may vary in intensity. Ventral surface of thigh may be pinkish. White dots outlining dorsal blotches may vary in size and number, forming distinct lines in some individuals; these white dots/lines may be greenish in live individuals. Few individuals have dorsal pattern weakly distinct in life and in preservative. Dorsal arrow-shaped blotch may present interruptions and be slightly irregularly shaped. Irregularly shaped small blotches scattered across dorsal surfaces of head and body may vary in number, size, shape, position, and distinctness from the background. Background color of belly may be bluish in different intensities. Brown coloration of the ventral border of mandible is more visible in fixed specimens, after the yellow coloration has faded. In fixed specimens it varies from occupying just the edge of the mandible to the anterior half of gular region. Tympanum may be slightly discernible externally in some individuals. Snout may be rounded in lateral view. Dorsum of the head, body, thighs, and tibiae may be slightly rugose. Dark superficial keratinized layer of the nuptial pad may be peeled; however, nuptial pad remains visible.
Advertisement and aggressive call
We identified two call types, the advertisement and an aggressive call (of undetermined specific function, see Toledo et al. 2015). Both the advertisement (n = 386 calls from seven individuals) and aggressive calls (n = 43 calls from four individuals) presented a pulsed structure. They differentiate from each other mainly by the call duration, note rate, and the context in which the recordings were obtained (advertisement calls recorded in the field and aggressive calls recorded from the aqua-terrarium). The advertisement calls had variable number of notes (1–3), frequency, and amplitude of notes varying in the call ( Fig. 4 View Fig ; Table 2 View Table 2 ). These calls seemed to be composed of a harmonic structure, but the harmonics were not clear in most of the calls. The most common advertisement call structure was that with 3 notes (exhibited in 63.63% of calls, 34.1% had two notes and 2.27% had only one note) with the dominant frequency in the second note (48.64% of the calls, 35.13% of calls had the dominant frequency in the third note and 16.21% in the first note). This call was emitted at rate of 0.87–2.04 calls/second, lasting between 0.07 to 0.3 seconds, with a dominant frequency of 0.94–2.63 kHz (average of 1.7 kHz ± 0.31 SD). On the other hand, we analyzed 1306 aggressive call notes from four males ( Fig. 5 View Fig ; Table 3 View Table 3 ). These calls were emitted by males close to other calling males, but it was not possible to determine a specific function (e.g., territorial, encounter or fighting calls). Aggressive calls were organized in groups of 9–118 notes, lasting 4.73–53.46 seconds, longer than advertisement calls, and with a dominant frequency of 1.59–2.30 kHz.
Tadpole
Maximum total length 28.2 mm, at stage 37. Body depressed (BH/BW = 0.73–0.85; Fig. 6A–B, H View Fig ), 0.39–0.43 times TL; in dorsal view, ovoid with well-marked lateral constrictions at the spiracle level; in lateral view, ventral contour convex in gular and abdominal regions, with a well-defined constriction slightly anterior to the spiracle level. Snout truncated in lateral view and obtuse in dorsal view (BWN/ BWE = 0.71–0.75). Nostrils elliptical, small (ND/BL = 0.02–0.02), dorsally located, anterodorsally directed, slightly closer to the tip of snout than to the eyes (NSD/ESD = 0.38–0.44); presence of a continuous and elevated marginal rim, with a small fleshy projection on the medial portion ( Fig. 6E View Fig ). Eyes dorsally located (IOD/BWE = 0.55–0.64), dorsolaterally directed, 0.20–0.25 times BWE. Spiracle sinistral, lateral, visible in dorsal and ventral views (SVD/BH = 0.47–0.60), posterodorsally directed, short (SL/BL = 0.08–0.10), opening at the middle third of the body (SSD/BL = 0.55–0.58); inner wall fused to the body, with its distal portion as a slight ridge ( Fig. 6F View Fig ). Vent tube dextral, posteriorly directed, short (VTL/BL = 0.07–0.10), with a large opening, fused to the ventral fin, and positioned at its ventral margin ( Fig. 6G View Fig ). Tail moderately high, with about the same height as the body (MTH/BH = 0.95–1.00); tail musculature slender (TMH/BH = 0.29–0.39), straight, not reaching the broadly rounded tip of tail. Dorsal and ventral fins about the same height (DFH/TAL = 0.11–0.14; VFH/TAL = 0.11–0.14), with the external margins slightly convex. Dorsal fin emerging on the posterior third of the body at a moderate sloping (DFIA = 15–17°); maximum height at the middle third of the tail. Oral disc medium-sized (ODW/ BW = 0.31–0.35, measured with oral disc closed), anteroventrally positioned (ODP = 33–43°), laterally emarginated ( Fig. 6D View Fig ); single row of conical and alternate marginal papillae interrupted anteriorly by a wide anterior gap (AGL/ODW = 0.58–0.60); few (3–4) small submarginal papillae arranged in a small row at the supra-angular region and other (1–3) aligned at the fold of the oral disc emargination. Labial tooth row formula (LTRF) 2(2)/3(1); A2, frequently irregular at the lateral portions, longer than A1, which is irregular along its length; P1 and P2 equal in length, slightly longer than P3; jaw sheaths wide, finely serrated on the margins (about 36 serrations on the upper sheath), upper jaw sheath M-shaped and lower jaw sheath V-shaped. Stitches of lateral line system not distinct; nerves of the ventral body-line and longitudinal oral line evident laterally, and nerves of dorsal and middle lines in the posterior portion of body. Intestinal tube circularly coiled ( Fig. 6C View Fig ), switchback point slightly dislocated from the center of the abdominal region. Measurements are shown in Table 4 View Table 4 .
Tadpole coloration
In preservative, body densely covered by dark brown melanophores, except the gular region, which is pale ( Fig. 6A–C View Fig ); intestine tube barely visible, almost covered by melanophores; rectus abdominis visible from the region of posterior limbs to the peribranchial region; distal portion of spiracle not pigmented. Tail musculature cream, homogeneously pigmented by melanophores; fins translucent, finely reticulated with filiform melanophores mainly the dorsal fin.
In life, body dark brown, finely speckled with iridophores ( Fig. 6H View Fig ); spiracle translucent; venter cream in the gular and abdominal regions; iris black with golden dots scattered and a narrow golden rim surrounding the pupil. Tail musculature cream, homogeneously covered with melanophores except by few small, depigmented areas; fins translucent, finely reticulated with filiform melanophores and golden dots; dorsal fin more pigmented than ventral fin.
Karyotype
The diploid chromosomal complement of P. araxa sp. nov. is composed of 22 chromosomes, with 8 pairs of metacentric chromosomes (1, 2, 5, 6, 7, 8, 9, and 10; pairs 2 and 7 are at the threshold between metacentric and submetacentric classification), 2 pairs of submetacentric chromosomes (3 and 4), and a telocentric chromosome pair (11) ( Fig. 7 View Fig ; Table 5 View Table 5 ). Large amounts of C-banded constitutive heterochromatin were detected in the centromeric region of all chromosomes. Faint C-bands were also observed interstitially on the long arm of chromosome 3 in some metaphases (inset in Fig. 7B View Fig ).
Phylogenetic inferences and genetic distances
In both MrBayes and TNT analyses, the new species, together with Physalaemus cf. araxa from Santa Teresa, ES, composed a highly supported clade that was the sister group of P. maculiventris in the Physalaemus signifer clade ( Fig. 8 View Fig ). Two genetic lineages were recognized in the P. maculiventris clade, one represented by the specimens from Bananal, SP, and another by specimens from three other localities ( Fig. 8 View Fig ). High genetic distances were found between these P. maculiventris lineages and also between each of them and the new species ( Table 6 View Table 6 ). In contrast, the p -distances estimated from the 16Sar-16Sbr and H1 fragments, between the specimen from Santa Teresa and the new species, were low, 1.48 and 2.57, respectively ( Table 6 View Table 6 ).
Remarks
The phylogenies presented in Lourenço et al. (2015) and in Leal et al. (2020) have a terminal named “ P. signifer (Congonhas do Campo, MG)”. The city name “Congonhas do Campo” was incorrectly used as it has changed to “Congonhas” since 1948. Herein we fixed it, changing the terminal name to “ P. signifer (Congonhas, MG) ” to avoid any misunderstandings regarding the city name.
Natural history
Individuals of Physalaemus araxa sp. nov. reproduce aggregated in shallow temporary swampy ponds ( Fig. 9 View Fig ) in altitudinal grasslands (campo de altitude) with interspersed granitic outcrops, a typical phytophysiognomy of the highlands of the Mantiqueira mountain range ( Safford 2007; Vasconcelos 2010). Individuals were registered between 2551 m a.s.l (Lagoa da Sombra) and 2656 m a.s.l. (Três Lagoas), P. araxa sp. nov. was the only anuran species found breeding at those high-altitude ponds (observation also corroborated by the autonomous recordings). After amplexus, pairs produced floating foam nests where eggs were laid. Two foam nests were collected on 5 October 2017. Ten days after, tadpoles started leaving the nests and at 12 November tadpoles reached the stage 25. We counted 187 tadpoles from those nests. Benthic exotrophic tadpoles (ecomorphological guild II: A:1 sensu Altig & McDiarmid 1999) were observed in the wild in October and December 2017. Calling activity, foam nests, and tadpoles were registered between October 2017 and December 2017. Immediately metamorphosed froglets (with no evidence of tail), raised in captivity, had between 7.76 and 8.85 mm (ZUEC-AMP 24403–5).
Males were found calling from the ground or on top of peat moss ( Sphagnum sp. ) hidden on the pond’s margins. During the complete day (24 h) that we recorded P. araxa sp. nov. vocal activity ( Fig. 10 View Fig ), males called during all 24 h, however, clearly presenting predominant nocturnal calling activity. Most of the calls were recorded from 18:00 to 3:00, reaching the peak of calling activity between 19:00 and 21:00.




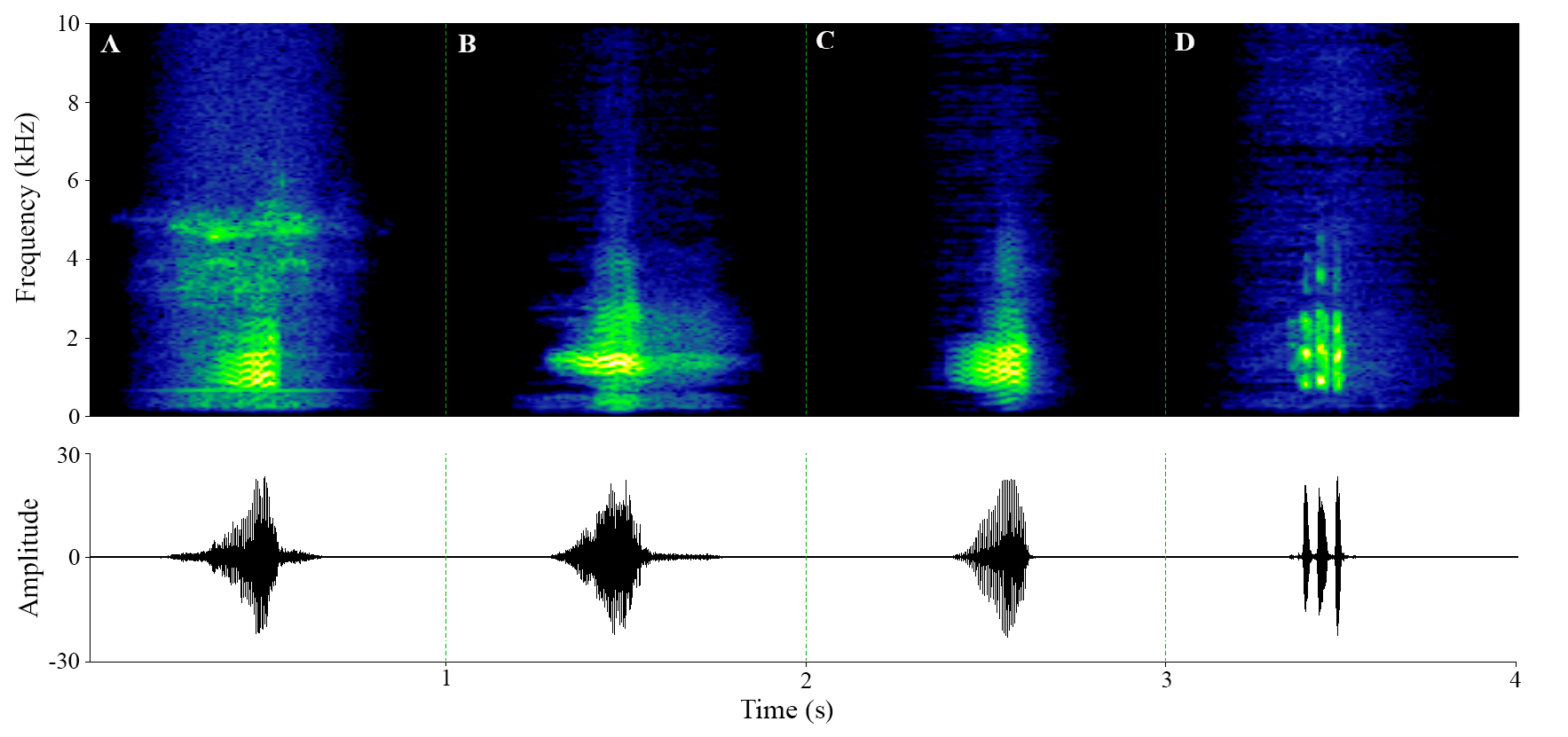
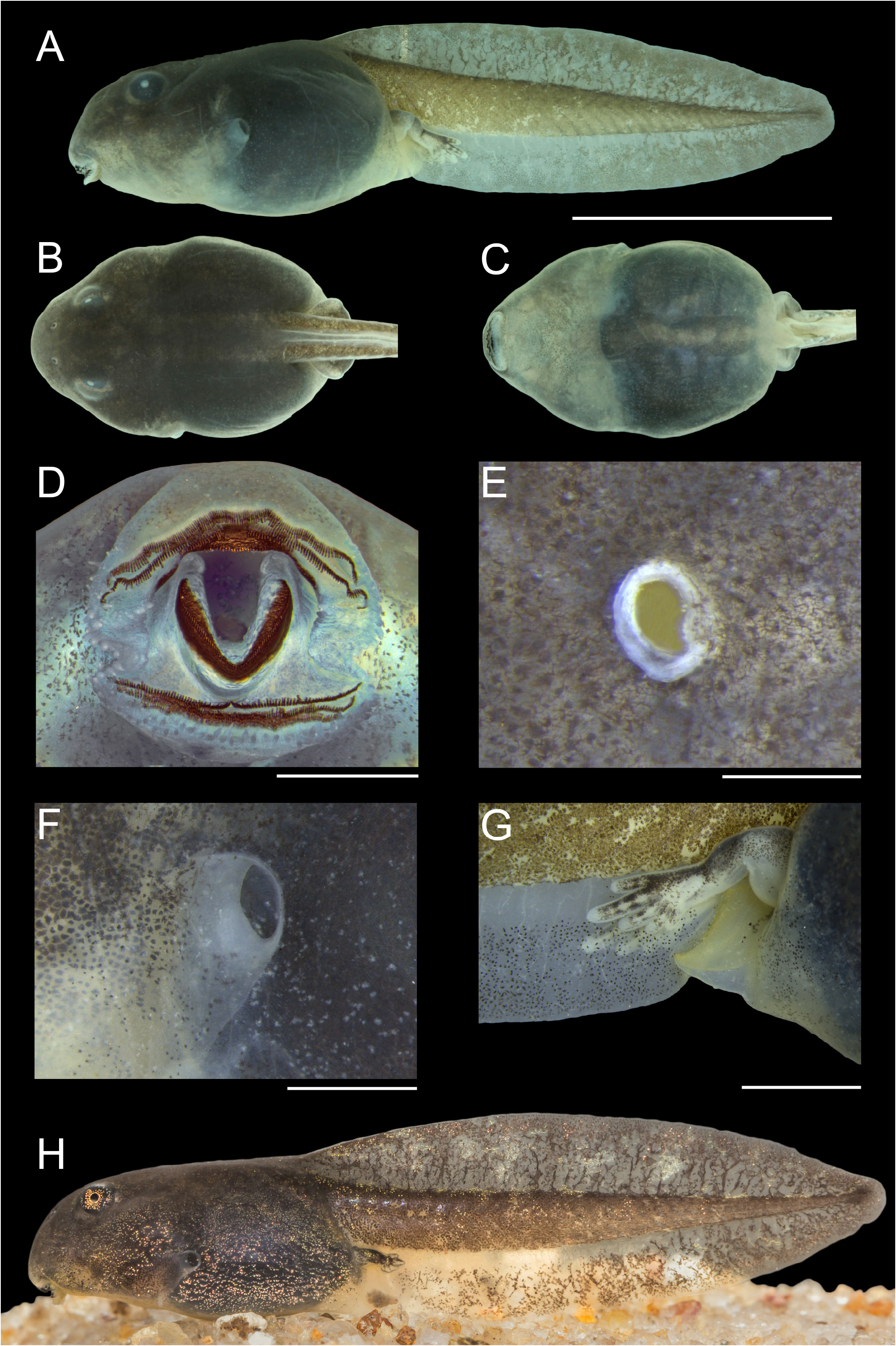
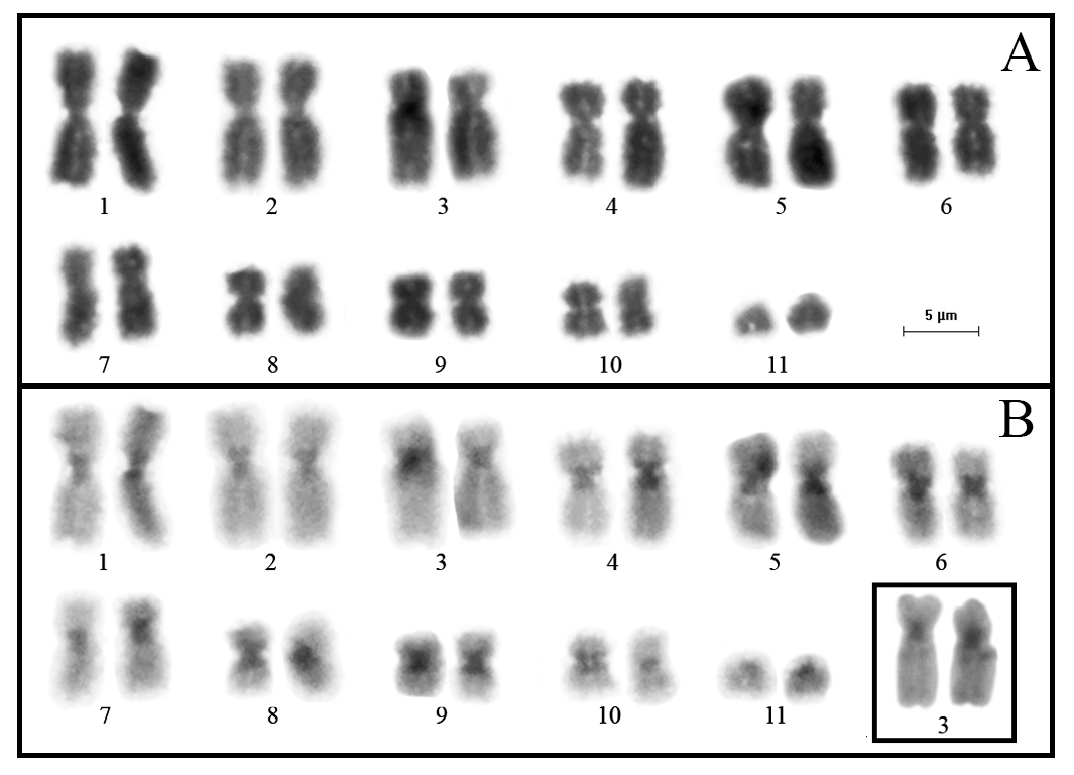



Table 4. Measurements (in mm) and angles of tadpoles of Physalaemus araxa sp. nov. (n = 9; ZUEC- AMP 24214) for the stages 37–38 (Gosner 1960). Data are presented as range (mean ± SD). See Material and methods for abbreviations.
| Measurement | Physalaemus araxa sp. nov. |
|---|---|
| TL | 25.2–28.2 (26.9 ± 1.1) |
| BL | 10.6–11.5 (11.1 ± 0.4) |
| TAL | 14.6–16.9 (15.8 ± 0.8) |
| MTH | 5.2–6.1 (5.7 ± 0.3) |
| DFH | 1.7–2.2 (2 ± 0.2) |
| VFH | 1.8–2.2 (2 ± 0.2) |
| TMH | 1.7–2.2 (1.9 ± 0.2) |
| BH | 5.5–6.1 (5.8 ± 0.3) |
| SL | 1–1.1 (1 ± 0.1) |
| SVD | 2.7–3.3 (3 ± 0.3) |
| SSD | 6–6.6 (6.3 ± 0.3) |
| ED | 1.1–1.4 (1.2 ± 0.1) |
| BW | 7.2–7.6 (7.4 ± 0.2) |
| BWN | 3.3–3.8 (3.6 ± 0.2) |
| BWE | 5–5.8 (5.4 ± 0.3) |
| TMW | 1.4–1.9 (1.7 ± 0.2) |
| END | 1.7–1.8 (1.8 ± 0.1) |
| ESD | 2.7–3.1 (2.9 ± 0.2) |
| NSD | 1.1–1.4 (1.2 ± 0.1) |
| ND | 0.2–0.3 (0.2 ± 0) |
| IND | 1.1–1.2 (1.2 ± 0) |
| IOD | 3.1–3.4 (3.2 ± 0.1) |
| ODW | 2.3–2.6 (2.4 ± 0.1) |
| ODP | 30–43.3 (34.4 ± 4.7) |
| AGL | 1.4–1.5 (1.5 ± 0) |
| DFIA | 15–17 (16.2 ± 1) |
| VTL | 0.8–1.4 (1.1 ± 0.2) |
Table 6. Uncorrected p-distances (%) between Physalaemus araxa sp. nov., P. maculiventris, and Physalaemus cf. araxa from Santa Teresa, ES. In the lower triangle, values estimated from H1 fragments; in the upper triangle, values inferred from the 16Sar-16Sbr fragment of the 16S rRNA gene (540 bp). Diagonal line presents uncorrected p-distances identified within each species or lineage, based on the H1 (left) and 16Sar-16Sbr (right) fragments. En-dash denotes that only one sequence is available.
| 1 | 2 | 3 | 4 | |
|---|---|---|---|---|
| 1. Physalaemus araxa sp. nov. | –/– | 1.48 | 5.83 | 5.20 |
| 2. Physalaemus cf. araxa – Santa Teresa | 2.57 | –/– | 6.94 | 6.12 |
| 3. P. maculiventris (topotype included) | 5.89 | 6.26 | 2.0/2.0 | 3.72 |
| 4. P. maculiventris – Bananal | 6.39 | 6.59 | 3.31 | 0.4/0 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Leiuperinae |
|
Genus |
Physalaemus araxa
| Leal, Fernando, Zornosa-Torres, Camila, Augusto-Alves, Guilherme, Dena, Simone, Pezzuti, Tiago Leite, Leite, Felipe, Lourenço, Luciana Bolsoni, Garcia, Paulo & Toledo, Luís Felipe 2021 |
Physalaemus sp.
| Zornosa-Torres C. & Augusto-Alves G. & Lyra M. L. & Silva J. C. & Garcia P. C. A. & Leite F. & Verdade V. & Rodrigues M. T. & Gasparini J. L. & Haddad C. F. B. & Toledo L. F. 2020: 4 |