
Antiquis opaque, 2014
|
publication ID |
https://doi.org/10.1111/zoj.12149 |
|
persistent identifier |
https://treatment.plazi.org/id/0388726A-4476-FF8F-679E-3985FAE0FE12 |
|
treatment provided by |
Marcus |
|
scientific name |
Antiquis opaque |
| status |
sp. nov. |
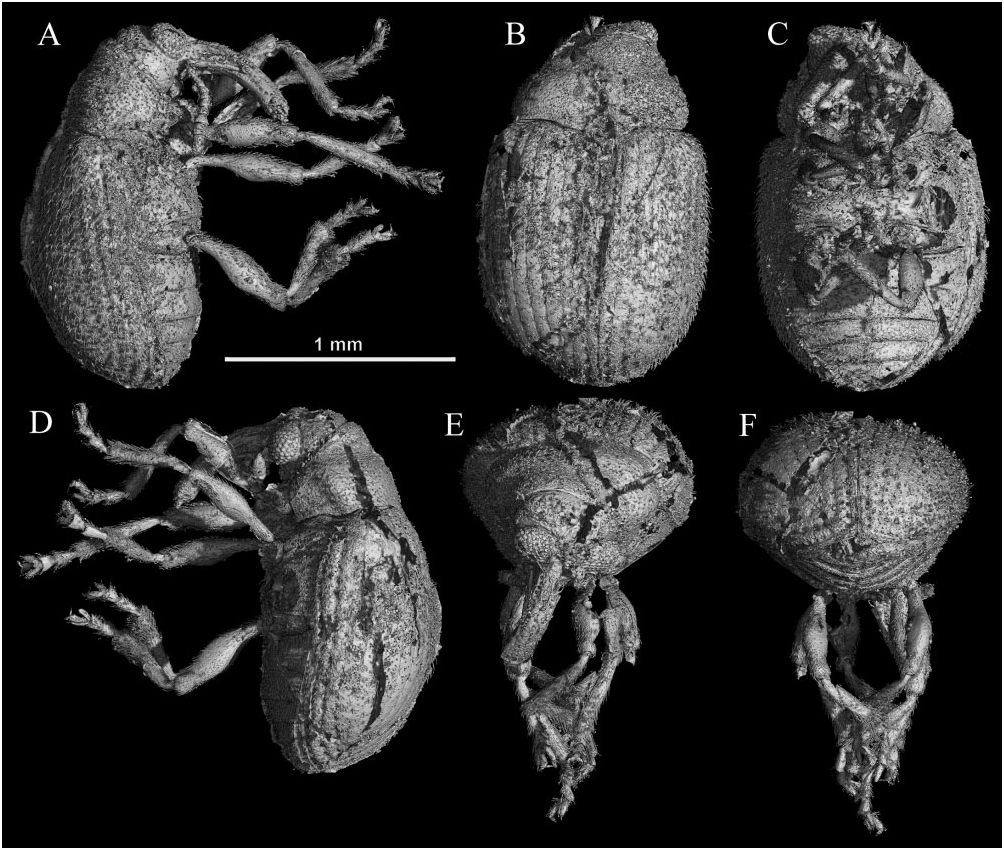
ANTIQUIS OPAQUE PERIS, DAVIS, ENGEL ET DELCLÒS SP. NOV. ( FIG. 5 View Figure 5 )
Etymology
The specific epithet is the noun opaque , and was selected for two reasons, i.e. the opaque amber in which the fossil is embedded and the rather confusing suite of characters that it exhibits. The term is treated as a noun in apposition.
Holotype
IGR.ARC–331.2, housed in the amber collection of the Geological Department and Museum of the University of Rennes 1 ( France). It is a complete specimen preserved in a piece of fully opaque amber, imaged by synchrotron microtomography. In the reconstruction it is possible to observe some cracks along the body, one in the right lateral side and one dorsally. The holotype was initially preserved as a syninclusion with one spider ( Lagonomegopidae ), two beetles ( Artematopodidae and one undetermined larva), and one crustacean (Tanaidacea).
Type locality and horizon
Font-de-Benon quarry, c. 1 km east of Archingeay (Charente-Maritime, France). The piece was found in the lithological level A1sl–A sensu Perrichot et al. (2010) [= A1sl1 sensu Néraudeau et al., (2002)]; mid-Cretaceous, latest Albian, or earliest Cenomanian in age.
Diagnosis
As for the genus ( vide supra).
Description
Body length 1.59 mm (without rostrum), maximum body width 1.00 mm, ratio of body length to greatest width 1.59; body ovate, pear-shaped, dorsoventrally very convex with margins rounded, oval in section in mesothorax; body covered by slightly appressed, dense pubescence.
Head transverse, same size as anterior part of pronotum, not constricted behind compound eyes; compound eyes large, rounded, rather coarsely faceted, very convex and protruding, posterior margins somewhat more produced; narrowly separated, with forehead narrower than rostrum width ( Fig. 5A, B View Figure 5 ). Rostrum broadly curved, slightly longer than pronotum, subquadrate in cross-section ( Fig. 5E View Figure 5 ); from base, rostrum length 0.51 mm, 1.3 times length of prothorax. Mouthparts mostly obscured, but three-segmented maxillary palpus and endodontous mandible visible. Antennae inserted laterally at middle of rostrum; antennae reaching to approximate middle of prothorax when directed posteriorly. Antennae with 11 antennomeres, geniculate; funicle with six articles (including pedicel) and apical compact club formed by four antennomeres, although the small apical fourth antennomere is difficult to distinguish in the reconstruction and may be misinterpreted ( Fig. 5A, D View Figure 5 ). Scape forming longest article, similar in length to remainder of funicle, narrower basally and wider apically, approaching anterior margin of compound eye but not touching it; pedicel (funicular article 1) similar in length to remainder of funicle; each funicular article 0.3 times length of scape; funicular articles 3 to 6 transverse, somewhat compact, gradually becoming wider towards apical articles, 0.6 times length of pedicel; club slightly compacted, with four articles. Club article 1 1.1 times length of funicular article 6 but 1.3 times wider; club article 2 similar in width but 1.3 times as long as preceding article; club article 3 0.7 times length of preceding article and narrowing apically; terminal club article 4 pointed and short ( Fig. 5D View Figure 5 ), 0.6 times length of preceding article; antennal scrobe faintly present, shallow.
Prothorax length 0.32 mm, maximum width 0.62 m; prothorax transverse, at apex 0.8 times as wide as length, in middle 1.6 times as wide as length, at base 1.9 times as wide as length, convex, narrower anteriorly than posteriorly; pronotal lateral margins sinuate, rounded; base of prothorax slightly narrower than elytra; sinuate, with medial part slightly emarginate and rounded ( Fig. 5B View Figure 5 ). Prosternum very short, only slightly longer than one procoxal diameter; procoxal cavities narrowly separated by less than one coxal diameter; procoxae conical and protruding ( Fig. 5C View Figure 5 ). Mesocoxae more widely separated by approximately one coxal diameter, mesocoxae circular and slightly protruding. Metacoxae horizontally orientated, separated by similar length as mesocoxae; metacoxae extending laterally, but not reaching elytra, completely separated from metaventrite by lateral suture of metaventrite.
Mesoscutellum small, transverse, 2.5 times wider than long. Elytral length 0.81 mm, 1.2 times as long as wide at humeri, 1.1 times as long and wide in middle, 1.5 times wide at apical quarter, width coinciding with maximum width of body; elytra very convex, lateral margins subparallel, very slightly widening medially, broadly curved and strongly rounded apically; ten punctate striae on each elytron ( Fig. 5A, B, F View Figure 5 ).
All legs similar in length ( Fig. 5A, D View Figure 5 ), covered by conspicuous, fairly dense setae; trochanters short, oblique; femora touching coxae; femora wider at middle; tibiae similar in length as femora, with a brush of dense, rigid setae along apical half; apical comb of setae transverse; tibiae lacking a mucro or uncus at apical margin; tibial spurs absent. Tarsomere 1 widening and emarginate apically; tarsomere 2 shorter than tarsomere 1 and more strongly emarginate; tarsomere 3 strongly bilobed and 1.2 times length of preceding tarsomere; tarsomere 4 extremely short and hidden between lobes of tarsomere 3; tarsomere 5 1.1 times length of tarsomere 3; pretarsal claws divergent, appendiculate with a basal swelling on each claw.
Abdominal ventrites separated by straight sutures ( Fig. 5C View Figure 5 ); ventrite 1 longest, 1.3 times as long as ventrite 2, with anteromedial process subacute, posteromedial margin broadly concave; ventrites 1 to 4 decreasing in width and length, ventrite 2 1.3 times as long as ventrite 3, ventrite 3 1.2 times as long as ventrite 4; ventrites 4 and 5 similar in length, ventrite 5 much narrower than 4 and strongly rounded apically. All tergites covered by elytra.
Remarks
The general habitus appears similar to Caridae and Nanophyinae (Brentidae) . The somewhat compact antennal funicle and club, the short, oblique trochanters, the straight sutures of the abdominal ventrites, and the proportionately long ventrites 3 and 4 do not agree with Brentidae . The geniculate antennae, compact antennal funicle and club, widened pronotum, and apparent lack of tibial spurs excludes the genus from Caridae ( sensu Oberprieler et al., 2007) . Although species of Car and several fossil taxa belonging to Caridae appear to have geniculate antennae (e.g. Cretonanophyes Zherikhin, 1977 ), this articulation between the scape and pedicel is not the derived form as present in Curculionidae (and similar in Nanophyinae and some Apioninae ), whereby the articulation is truly in a lateral orientation. Antiquis gen. nov. is therefore placed in Curculionidae based on the truly geniculate antennae, compact antennal funicle and club, and short, oblique trochanters. Although this specimen does not possess any apical tibial modifications, such as an uncus or mucro, which are fairly typical in Curculionidae , such inermous tibiae are not entirely uncommon across this lineage, e.g. representatives of Entiminae , Curculioninae , and Erirhininae ( sensu Oberprieler et al., 2007) .
Some members within Erirhininae appear strikingly similar to this specimen, particularly with regard to the antennal funicle of six articles, rather short pronotum, procoxae that are positioned in the middle of the prosternum, and rather short fifth tarsomeres. Although this group tends to share many external features with other curculionids (particularly Curculioninae ), leaving the majority of its synapomorphies in genital characteristics, several features of erirhinines seem to exclude an inclusive placement of this specimen. These features include comparatively much shorter third and fourth ventrites, antennae nearly always inserted subapically on the rostrum, tibiae typically apically armed with an uncus and/or mucron, a narrow and parallel-sided pronotum, and slightly umbonate subapical area on the elytra. Although such external features may be considered subtle and fairly plastic within Curculionidae , this specimen possesses a combination of characters apparently not present in Erirhininae .
Anderson (2002) describes members of Curculioninae as (1) possessing an apical angle of the tibiae without spines, (2) compound eyes rounded, (3) a long rostrum, which is cylindrical in cross-section, and (4) the scape not reaching the anterior margin of the compound eye. This fossil specimen demonstrates all of these features, with the exception of the cylindrical rostrum, for which the fossil bears a more subquadrate rostrum in cross-section. The fossil has a particularly close resemblance to members of the tribes Derelomini ( sensu Franz, 2006) and Anthonomini, which may also demonstrate protuberant compound eyes, antennal funicle of six articles, tibiae with transverse apical setal combs, inermous tibial apices (lacking any modifications such as an uncus or mucro), divergent (as well as appendiculate) pretarsal claws, abdominal ventrites that become proportionately shorter from the first to fifth (i.e. not with ventrites three and four much shorter than the others), and broadly smooth elytra lacking any umbonate regions.
Although this resemblance is strong, the fossil is clearly primitive within the subfamily owing to the somewhat incomplete compaction of the antennal club. It then seems fairly justified, as well as conservative, to place this specimen in Curculioninae but as tribe incertae sedis pending the discovery of additional material and its inclusion in a formal cladistic analysis.
Two fossil weevils from the earliest Cenomanian Burmese amber were described by Poinar (2006) and Poinar & Brown (2009). Palaeocryptorhynchus burmanus Poinar, 2006 , is definitively placed in Curculionidae based on the pronotum extended overhead, toothed femora, and tibiae with a terminal, simple uncus; none of these characters was observed in Antiquis gen. nov. However, Anchineus dolichobothris Poinar et Brown, 2009 , generally has some resemblance to the new French specimen. Even so, Anchineus Poinar et Brown, 2009 , can be distinguished clearly from Antiquis gen. nov. by its funicle of seven articles, club of three articles, and bifid pretarsal claws; whereas Antiquis gen. nov. has a funicle of six articles, club of four articles, and pretarsal claws with only a basal swelling. Two other curculionids are described from Lebanese and Burmese ambers; however, they belong to Scolytinae and could not be confused with Antiquis gen. nov. Apparently, some other basal curculionid lineages appear as compression fossils from the Aptian (Early Cretaceous) of Bon- Tsagaan ( Mongolia), although the precise identification and placement of these are somewhat questionable and should be interpreted cautiously because of their fairly incomplete preservation ( Legalov, 2012b).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.