
Phyllomedusa chaparroi, Castroviejo-Fisher, Santiago, Köhler, Jörn, Riva, Ignacio De La & Padial, José M., 2017
|
publication ID |
https://doi.org/10.11646/zootaxa.4269.2.4 |
|
publication LSID |
lsid:zoobank.org:pub:79C7F208-9952-48FA-AEEB-679B94D86240 |
|
DOI |
https://doi.org/10.5281/zenodo.6003982 |
|
persistent identifier |
https://treatment.plazi.org/id/038987D6-1600-FFBE-FF33-F9437EB788E1 |
|
treatment provided by |
Plazi |
|
scientific name |
Phyllomedusa chaparroi |
| status |
sp. nov. |
Phyllomedusa chaparroi sp. nov.
Holotype. MUBI 13986 (field number AJC 2613), adult male, from Shucshuyacu , 06°37'20.7'' S, 76°36'53.9'' W, 537 m above sea level, Departamento San Martín, Peru, collected on 25 March 2010 by S. Castroviejo-Fisher. GoogleMaps
Paratypes. MNCN 46650, 46651 (field numbers AJC 2682, 2683) and MUBI 13987, 13988 (field numbers AJC 2684, 2685), four adult males, from the road from Tarapoto to Tocachi, 07°41'26.5'' S, 76°39'53.9'' W, 650 m a.s.l., Departamento San Martín, Peru, collected on 26 March 2010 by S. Castroviejo-Fisher and M. Guerra Panijo. GoogleMaps
Generic placement. A species assigned to the genus Phyllomedusa based on phylogenetic analyses and presence of parotoid glands and toe I longer than toe II. We consider P. chaparroi a member of the P. tarsius species group ( sensu this work, see Discussion) based on our phylogenetic analysis of combined DNA sequences of mitochondrial and nuclear genes (Fig. 2) and because it shares a unique iris coloration with species of that group (see Discussion, Fig. 5 View FIGURE 5 ).
Diagnosis. (1) moderate size (adult males SVL 67.9–77.5 mm); (2) palpebral membrane transparent, not reticulated; (3) in life, dark reddish-brown iris without distinct reticulation; (4) vomerine odontophores present, bearing teeth; (5) parotoid glands present in sexually active males; (6) dorsal skin coarsely shagreen, lacking keratinized tubercles ( Fig. 3 View FIGURE 3 D); (7) first toe longer than second; (8) low dermal ridges on outer edge of forearm and tarsus, extending on outer edges of fourth finger and fifth toe, respectively; (9) in life, dorsal surfaces green; upper eyelid, upper lip and concealed surfaces of limbs and flanks yellow; ventral surfaces creamy-white ( Fig. 3 View FIGURE 3 A–C).
Comparisons. (Characters of the new species are in parentheses.) Species in the Phyllomedusa tarsius group ( sensu Barrio-Amorós 2006, P. coelestis , P. neildi , P. tarsius , P. trinitatis , and P. venusta ) have a distinct iris reticulation in life (dark reddish-brown iris without obvious reticulation, see Fig. 5 View FIGURE 5 E–H).
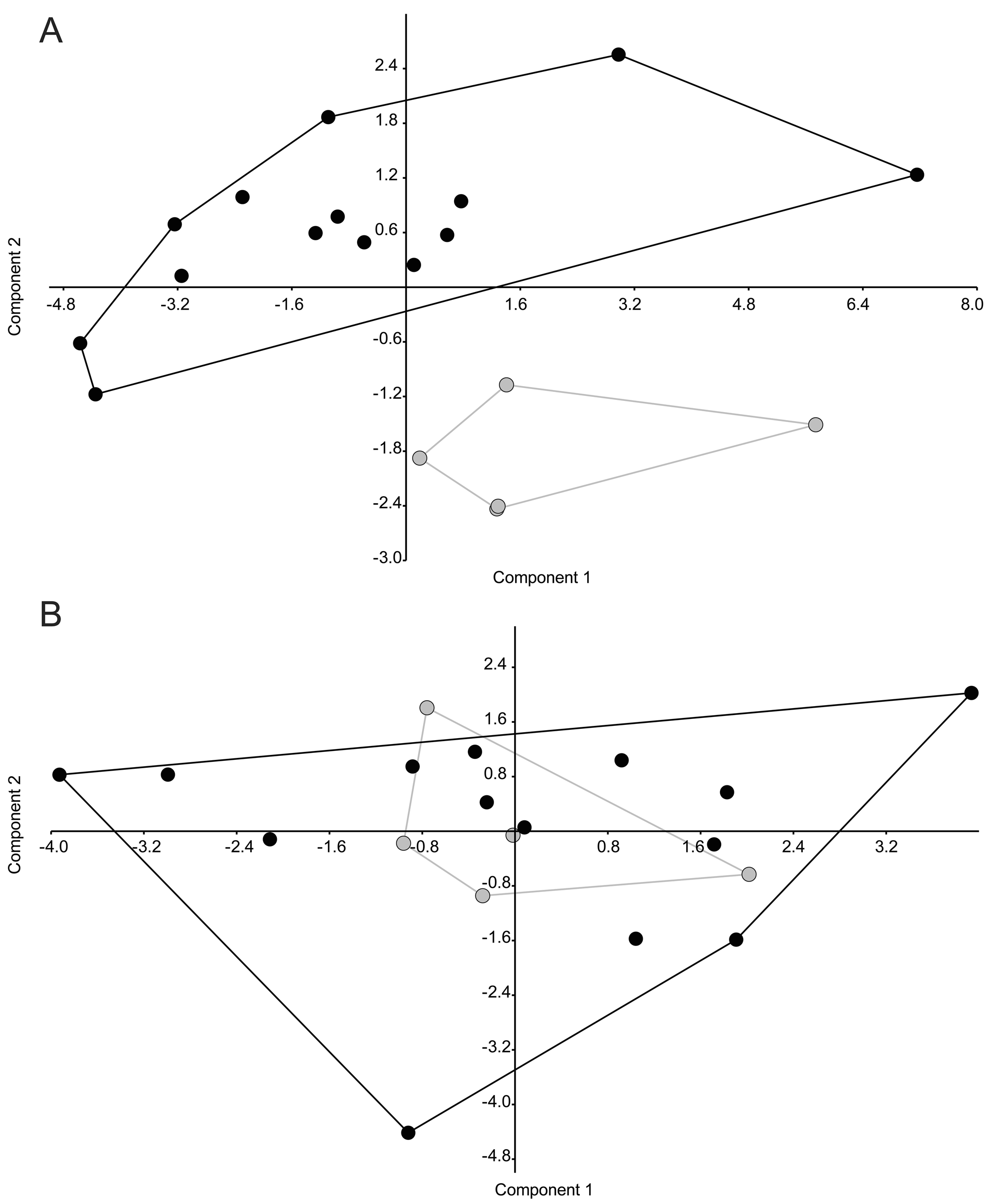
In external morphology, the new species is most similar to P. boliviana and P. camba (both currently not included in any species group). Phyllomedusa boliviana has a red upper eyelid rim (yellow), a creamy white upper lip (yellow), cream flanks, sometimes with flat reddish tubercles or markings in life (yellow), and a black iris in life (dark reddish-brown). We were not able to detect any qualitative morphological character distinguishing the new species from P. camba . The PCA analysis of the raw measurements of males of P. camba and P. chaparroi ( Fig. 1 View FIGURE 1 A) show no overlap in morpho-space, but when corrected for the effect of body size (SVL), variation in measurements of both species completely overlap ( Fig. 1 View FIGURE 1 B) and the first two components only explained 43.6 % of the variance. Thus, morphometric differences among specimens of both species seem to be explained by overall size, which is likely dependent on age and environmental conditions ( e.g., availability of food), and should not be used as evidence of species divergence. We relied on phylogenetic position to justify the recognition of the new species when compared to P. camba , as our phylogenetic analyses (both MP and ML) indicate that P. chaparroi is more closely related to species of the P. tarsius group ( sensu Barrio-Amorós 2006) than to P. boliviana or P. camba . Genetic distances between P. chaparroi and species of the P. tarsius group and P. camba range between 2.8–5.5 % ( Table 3). We identified 25 molecular transformations diagnosing P. chaparroi (Appendix 3).
Species in the Phyllomedusa burmeisteri species group ( P. bahiana , P. burmeisteri , P. distincta , P. iheringii , P. sauvagii , and P. tetraploidea ) lack a dark reddish iris (dark reddish-brown), and present distinct orange flecks on flanks— P. burmeisteri , P. distincta , P. iheringii —or blue flanks— P. burmeisteri and P. bahiana —(absent). Phyllomedusa bicolor (not assigned to a species group) is a large species with a SVL in adult males of 91.0–103.0 mm (SVL 67.9–77.5 mm), has a silver iris (dark reddish-brown), a dorsolateral line made of white spots surrounded by dark blackish-brown pigments (absent), and larger parotoid glands (smaller parotoid glands). Phyllomedusa vaillanti (not assigned to a species group) has a dorsolateral row of enlarged tubercles (absent), orange/black pattern on flanks (absent), and iris pale gray in life (dark reddish-brown).
Species of Callimedusa (the P. perinesos species group sensu Ron et al. 2013) have purple colors on hands, feet, and concealed surfaces of limbs, and a purple venter with white granules in life (purple colors absent in life). Callimedusa atelopoides shows a purple dorsum (green in the new species), is terrestrial (arboreal); adult males in C. tomopterna are 40.0–48.0 mm in SVL, present calcar tubercles, and orange-black striped pattern on flanks (males SVL 67.9–77.5 mm, calcars absent, and orange-black striped pattern on flanks absent). Additionally, species of Callimedusa do not have a reddish iris (dark reddish-brown).
Species of Pithecopus (the P. hypochondrialis species group sensu Faivovich et al. 2010) mainly differ from P. chaparroi by smaller size, maximum SVL of adult males < 44 mm (adult male SVL ± 67.9 mm), vomerine odontophores absent (present), iris not dark red or brown (dark reddish-brown), and orange marks in groin (orange marks absent).
Description of the holotype. An adult male of medium-size, right tarsus medially cut and preserved as tissue sample ( Fig. 4 View FIGURE 4 ; see Table 3 for measurements). Head about as wide as body width; snout short, rounded in dorsal view, inclined in lateral view; canthus rostralis slightly concave in dorsal view, rounded in profile; loreal region concave; lips thin and not flared; nostrils protuberant, directed laterally; internarial region flat; eyes protuberant; pupil vertically elliptical; palpebral membrane transparent, without reticulation, upper margin with a dark brown line; parotoid glands weakly developed, extending from eyelid to a level slightly posterior to insertion of forearm; supratympanic fold present, barely obscuring the upper edge of tympanum; tympanum distinct, oval, its anterior edge separated from eye by 2 mm; tongue large, suboval, posterior third free; vomerine odontophores present, prominent, located medially at midlevel between elliptical choanae, separated from each other medially by a distance equal to two thirds the width of one odontophore, each bearing five vomerine teeth; vocal slits short; vocal sac single, median, subgular, not distinct.
Upper arm slender (width 2.2 mm), forearm robust, wider (width 4.6 mm) than upper arm, with low lateral ridge extending to outer edge of fourth finger; ulnar fold distinct; relative length of fingers I<II<IV<III; finger tips about half the diameter of tympanum diameter; subarticular tubercles large, round to conical; supernumerary tubercles barely detectable; palmar tubercle oval, flat, indistinct; thenar tubercle elevated, elliptical; prepollex completely covered by skin; inner side of first finger bearing keratinized, dark brown nuptial callosities, covering about half of the length of first finger (terminal phalange not covered); palmar webbing absent.
Hind limb slender, moderately long, TL 44 % of SVL, without calcar or dermal ornamentation; inner tarsal fold absent; outer tarsal fold low, barely distinct; relative length of toes II<I<III<V<IV; toe tips slightly smaller than those of fingers; inner metatarsal tubercle elliptical, low, flattened; outer metatarsal tubercle indistinct, flat, ovoid; subarticular tubercles large, round to conical; no supernumerary tubercles on toes; plantar webbing absent.
Cloacal opening directed posteriorly at midlevel of thighs, unornamented; supra-cloacal fold present; skin of dorsal surfaces smooth in preservative (coarsely shagreen in life), tubercles absent; ventral skin, flanks, and inferior surfaces of thighs tubercular.
In life, dorsal surfaces and upper half of flanks (dorsum, head, tympanum, and upper parts of limbs) leaf green ( Fig. 3 View FIGURE 3 A, B). Dorsal surfaces of fingers and toes cream to yellowish. Border of upper eyelid lemmon yellow. Upper lip yellow from level of posterior edge of eye to tip of snout, creamy white below tympanum. Inner concealed surfaces of limbs lemon yellow. Flanks green with irregular lemon yellow flecks and markings, extending from insertion of forearm to level of thighs ( Fig. 1 View FIGURE 1 B); irregular white flecking at lower flanks. Dermal ridges on outer arm and tarsus whitish. Gular skin pinkish creamy white, with some irregular brownish marbling; two small rounded cream-colored flecks on gular region; a longitudinal row of three larger, more irregular cream flecks at central level of chest. Thin cream-colored supracloacal line. Venter creamy white with pinkish tint ( Fig. 3 View FIGURE 3 C). One rounded fleck on each side at ventral surfaces of thighs, close to cloaca. Palmar and plantar surfaces fleshy with a pinkish tint. Iris dark reddish-brown with fine irregular black flecking and tiny orange spots.
In preservative, same pattern as described above, but green dorsal surfaces faded to bluish-grey, with a violet tint ( Fig. 4 View FIGURE 4 ). Yellow colors on flanks and lips faded to cream with some pinkish tint. Throat and chest brown, with cream spots exhibiting pinkish tint. Posterior surfaces of thighs and region around cloaca brown with fine cream spotting.
Variation. For variation in measurements see Table 1. Generally, the four available male paratypes exhibit the same morphology and color pattern as the holotype. However, the parotoid glands are more prominent in the largest male (MUBI 13988) and posteriorly reach a point at a level where the elbow touches the body, when forelimb is adpressed posteriorly. Furthermore, MUBI 13988 exhibits a more contrasting coloration of the flanks, with well-defined cream-colored blotches, as well as few scattered elevated warts on the posterior part of the dorsum and additional cream flecking on the ventral surfaces of thighs. MUBI 13987 exhibits distinct cream spots on the posterior surfaces of thighs, lacking in all other specimens. In contrast to the holotype, all four paratypes exhibit a single cream fleck on chest, and lack additional flecks on the throat. However, this fleck on chest is divided by a thin brown line in MUBI 13987.
Natural history. All specimens were collected between dusk (19:00h) and midnight near temporary ponds on the side of roads. The vegetation surrounding the ponds was disturbed secondary forest, although in the vicinity there was primary or old grown secondary forest. Specimens were found active and perching on branches at 0.5– 1.5 m above the ground. The holotype was collected after a partially cloudy day with no rain. No vocal activity was observed. The paratypes were collected after a day of continuous rain. All of them were vocalizing (no recordings available) and to the human ear their calls superficially resembled those of Phyllomedusa tarsius from Amazonian Venezuela or P. camba .
Distribution. Although our type series only includes specimens from two sites, this species seems to occur at various localities in the area of Tarapoto in the Peruvian Amazon (E. Twomey and P. Venegas, personal communication). Nonetheless, it seems that this species has been confused with Phyllomedusa camba and the distributional range of both species needs to be thoroughly evaluated, especially at the northernwestern limits of the known distribution of P. camba ( i. e., the Peruvian departments of Madre de Dios and Ucayali, and the Brazilian states of Acre and Amazonas; see De la Riva 1999).
Etymology. The name is a patronym for Juan Carlos Chaparro Auza, a Peruvian herpetologist from Cusco, Peru, and is used as a noun in the genitive case. We name this species after our dear friend in recognition of his scientific contributions to the knowledge of the amphibians and reptiles of Peru and as a tribute to the good moments shared in the field.



| MNCN |
Museo Nacional de Ciencias Naturales |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.