
Petawrus norfolcensis (Kerr, 1792)
|
publication ID |
https://doi.org/10.5281/zenodo.6656820 |
|
DOI |
https://doi.org/10.5281/zenodo.6620353 |
|
persistent identifier |
https://treatment.plazi.org/id/038A1613-FFBB-FF90-83E3-F579C0F3F980 |
|
treatment provided by |
Felipe |
|
scientific name |
Petawrus norfolcensis |
| status |
|
12.
Squirrel Glider
French: Possum écureuil / German: Mittlerer Gleithornchenbeutler / Spanish: Falangero planeador mediano
Other common names: Squirrel Flying Phalanger
Taxonomy. Sciurus (Petaurus) norfolcensis Kerr, 1792 ,
Sydney, New South Wales, Australia.
This species is monotypic.
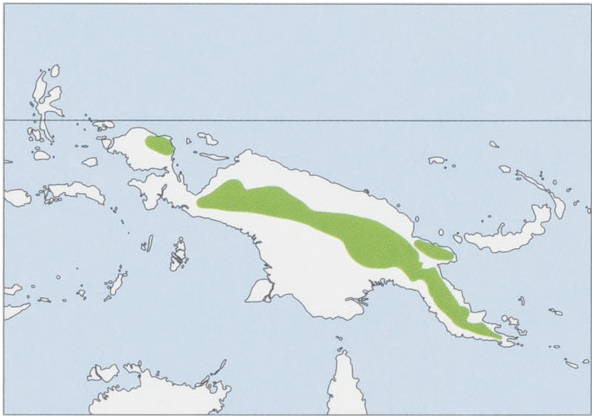
Distribution. E Australia from N Queensland (Cape York Peninsula) down E coast into C Victoria, including Ricketts and Fraser Is, also a few records in Western Flat and Bordertown, SE South Australia. View Figure
Descriptive notes. Head-body 18-24 cm, tail 22-30 cm; weight 173-300 g. The Squirrel Glider has soft and silky fur. Dorsal surface is a soft gray, face a paler gray, and there is a well-defined dark brown or black band commencing between eyes and running backwards along dorsal surface. Tail very bushy, especially basal third, where hairs often ¢.60 mm long. Ventral fur usually rich white, cream, yellow, or dark mustard. Similar in appearance to the Sugar Glider (P. breviceps), but larger, with longer and more pointed face, longer and narrower ears, and much bushiertail (especially at base); ventral fur does not have gray bases like that of the Sugar Glider. In north-east New South Wales and south-east Queensland, white ventral fur usually present only in juveniles.
Habitat. This glider occurs primarily in dry open woodland and forest where overstory includes myrtle family trees of genera Eucalyptus , Angophora , or Corymbia and understory includes trees of genera Acacia (Fabaceae) or Banksia (Proteaceae) . In south-east Queensland occurs mainly in areas containing plant species typical of drier eucalypt open forest and woodland;less frequently located in vegetation containing eucalypts and other plants typical of higher-rainfall areas, near rainforest, or along creeks and drain lines. In northern New South Wales occurs in dry forests and woodlands with overstory of winter-flowering eucalypts and understory of winter-flowering Banksia or Acacia . In Victoria it appears to be restricted to north-eastern riverine plains and uplands, and northern slopes of Western Highlands (Grampians), where it inhabits remnant woodlands or open forests that have mature or mixed-age stands of more than one eucalypt species, or riparian open forests of Eucalyptus camaldulensis. Trees and shrubs that provide a winter resource of nectar and hollow-bearing trees are apparently important components of habitat. Throughout range has typically been reported not above 300 m elevation.
Food and Feeding. Similar to other petaurid gliders,this species has a diverse diet that includes plant exudates as well as insects. Dietary items include nectar and pollen from various myrtaceous tree species such as Eucalyptus , Corymbia , Lophostemon , Melaleuca , and Angophora . Acacia gum and arils, the sap from eucalypts and invertebrates such as beetles, moth larvae, caterpillars, and spiders are also consumed. It also feeds upon lerps, honeydew, and lichen. A study of Squirrel Gliders in central Queensland found pollen of Eucalyptus / Corymbia in 70% and Melaleuca in 20% of fecal samples; also indications of sap-feeding in 44% of samples, seeds in 14%, and invertebrates (mostly moth larvae) present in 54% of samples. During this study, main food types were invertebrates (36% of observations), nectar and pollen (27% of observations), and sap (26% of observations) of five tree species. Although this speciesis typically an insectivore/exudivore, there are records of opportunistic predation of eggs of common bronzewing (Phaps chalcoptera), magpie-lark ( Grallina cyanoleuca ), and noisy miner ( Manorina melanocephala ). Studies of behavior of Squirrel Gliders in remnant forest in south-east Queensland revealed that feeding from flowers accounted for 48% of diet, with nectar and pollen derived from ten overstorey tree species; forest red gum (Fucalyptus tereticornis) dominated because ofits high abundance and protracted flowering period. Feeding on honeydew and lerp accounted for 15% and 2%, respectively, of all observations. Arthropods accounted for 35%, searched for in 20 different tree species, where a range of substrates was used; brushbox ( Lophostemon confertus) was the most important, being used in all seasons and accounting for 49% of these observations. These results contrast with assessments of Squirrel Glider diet at othersites, where a greater range of broad food types was exploited, and may reflect disturbed quality of habitat at the study site; nevertheless, they confirm importance of eucalypt nectar in the diet of this species.
Breeding. Breeding season is variable, but births have been recorded between March and December, with peak in April-July. All females more than one year old appear to breed each year. One or two young are born, mean litter size being c.1-7. Overall natality rate was 1-9 per year, indicating that females occasionally breed twice annually. The young typically disperse at 11-12 months of age and are capable of breeding at 12 months. Although typical longevity is thought to be 5-6 years, there is a record of an individual which lived to the age of seven years and nine months.
Activity patterns. Like other petaurid gliders, this species spends much of the night active. One study that explored its time budget revealed that it spent ¢.72% of the night in foraging, with 62% of that time devoted to feeding and 10% devoted to climbing and gliding. A further 16% of time was spent in grooming, though this may have been a response to being disturbed by humans. The remainder of the night was spent on other activities. The gliders rested (sitting stationary) for 5:8% of the time, engaged in social interactions with other gliders for 3-6%, and spent 2:5% of the time in their hollows.
Movements, Home range and Social organization. Squirrel Gliders have been found to nest in socially monogamous groups or mixed-sex groups of up to six individuals in leaf-lined tree hollows, primarily in species of eucalypt. They nest in dead trees, as well as in a variety ofliving trees, of Eucalyptus , Corymbia , Lophostemon , and occasionally Acacia . In different localities they can be monogamous or polygynous, groups tending to be composed of up to two males, of which only one is mature (more than three years of age), and two or more females and their offspring. They appear not to display any territoriality (though they appear to defend a core area). The home range is of 0-7-15 ha (typically between 2-9 ha and 8-8 ha), which they traverse in glides that average 20 m (range 9-47 m), though glides of 80 m have been recorded. Density estimates range from 0-35 ind/ha to 1-6 ind/ha, the number of animals present depending on food resources. One study showed a peak in density of c.1-6 ind/ha when there was significant nectar and pollen available during winter; density, however, declined to 0-5 ind/ha by fourth year, when there were extremely dry conditions and a lack of flowering in Eucalyptus siderophloia, an important nectar source. The gliders communicate with each other by using a range of calls. One of these is a monosyllabic or polysyllabic nasal grunt, ranging from a single call to sequences of up to 20 minutes’ duration (mean 2 minutes). Studies suggest that the rate of nasal grunts shows a positive relationship with population density and with proximity of conspecifics, but responses are infrequent; these grunts appear to regulate individual spacing by facilitating mutual avoidance. The other most common vocalizations are threatening calls that are accompanied by scuffles and/or chases. The preferred trees used for nesting in a south-east Queensland study were snags and iron-barked eucalypts. Even though non-eucalypt trees represented more than 9% of the available trees with diameters greater than 10 cm, no gliders that were successfully followed used them for nesting. A study of the den trees of Squirrel Gliders in south-east Queensland and north-east New South Wales found that the species used 13 species of living and dead tree across the two locations, dead trees accounting for a large percentage (more than 50%) of dens despite comprising only 3-10% oftrees with breast-height diameter greater than 20 cm at each location. Mean size of den trees at these sites was 49 cm in Queensland and 63 cm in New South Wales, den-entrance heights above ground averaging 7 m and 12 m, respectively; den entrances varied in width from 2-5 cm to 12 cm, with most less than 5 cm in diameter, and were most commonly located in fissures in the trunk and holes in branches. In observations of tree hollows utilized by Squirrel Gliders in south-eastern Australia, individuals used an average of seven hollow trees as den sites, these sites often adjacent to areas where nocturnal activities took place; average distance between den sites used by individual gliders on successive days was 218 m. Dens were often shared by an adult pair and a juvenile. Den trees were disproportionately used, gliders showing preference for one or two primary den trees often located on steep slopes.
Status and Conservation. Classified as Least Concern on The IUCN Red List. This species 1s, however, considered vulnerable in New South Wales, threatened in Victoria, and formally listed as endangered in South Australia, where it is known only from a skin collected near Bordertown (in upper south-east of South Australia) and as a result is generally considered to be extinct in that State. Across much ofits distribution there has been a steady loss of quality and extent of habitat owing to removal of timber for agriculture, sawn products, and firewood, and increasing coastal development, which results in clearance of forest remnants. Regeneration of trees and shrubs has also been hindered by grazing by stock, rabbits, and macropods, and by inappropriate fire regimes. In one study, microsatellite analysis was used to compare the functional (genetic) connectivity of different populations of Squirrel Gliders that have had different degrees of habitat fragmentation as a result of clearing since European settlement, which began in 1788. The extensive clearing for grazing (more than a century ago) of the species’ preferred habitat in south ofits distribution has meant that habitatis largely restricted to linear strips of remnant vegetation associated with roads, travellingstock reserves, and waterways. Other regions are more intact or have had more recent clearing. Populations with little, or at least recent, clearing had a greater genetic diversity than those where there had been a long history of clearing. Even some northern coastal populations associated with urbanization and agriculture showed evidence of genetic isolation. It is evident that the massive landscape changes following European settlement have had substantial impacts on levels of connectivity among Squirrel Glider populations (and no doubt those of other arboreal mammals). Landscape planning and management in the south should, therefore, be focused on restoring habitat connectivity where feasible, and along the coast existing habitat connectivity must be maintained and recent losses restored. On basis of the glide efficiency ofthis species, trees beside roads that create a tree-gap of 20 m (two-lane road) or 43 m (four-lane road) will need to be at least 13 m and 25 m, respectively, in height to enable animals to glide safely across them. Where trees are absent, wooden poles can potentially be installed to facilitate movement across roads and other barriers. Repeated use of such poles by Squirrel Gliders has been observed at several locations, suggesting that poles can help to restore habitat connectivity. Finally, an interesting situation regarding population dynamics was noted in a study in north-east New South Wales, Here, a remnant population of this species fluctuated considerably, and during a three-month period in winter-spring (July-September) rapidly lost weight, this coinciding with extremely dry conditions and a lack of flowering in Eucalyptus siderophloia, an important nectar source. The number of animals caught in the study grid fell by almost 80% between September and November; the total population declined by 55% and the adult population by 42% when compared with previous years. The number of gliders in the following year was low, suggesting that the population was slow to recover.
Bibliography. Ball et al. (2009), Beyer et al. (2008), Brearley, Bradley et al. (2011), Brearley, McAlpine et al. (2011), Carthew (2004), Crane et al. (2010), Dobson et al. (2005), Flannery (1994a), Goldingay & Taylor (2009), Goldingay, Sharpe, Beyer & Dobson (2006), Goldingay, Sharpe & Dobson (2010), Goldingay, Taylor & Ball (2011), Holland (2001), Holland etal. (2007), Jackson (1994), Malekian etal. (2006), Menkhorst & Collier (1987), Menkhorst et al. (1988), Millis & Bradley (2001), Quin (1995), Quin, Smith & Green (2004), Quin, Smith & Norton (1996), van der Ree (2002), van der Ree & Bennett (2003), van der Ree & Suckling (2008), van der Ree, Harper & Crane (2006), Rowston (1998), Rowston et al. (2002), Sharpe (2004), Sharpe & Goldingay (1998, 2007, 2009, 2010), Smith & Murray (2003), Taylor et al. (2011), Traill (1998), Traill & Lill (1997), Winter (1966), Winter, Lunney et al. (2008).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
SubClass |
Metatheria |
|
Order |
|
|
SubOrder |
Phalangeriformes |
|
SuperFamily |
Petauroidea |
|
Family |
|
|
Genus |
Petawrus norfolcensis
| Russell A. Mittermeier & Don E. Wilson 2015 |
Sciurus (Petaurus) norfolcensis
| Kerr 1792 |