
Bryoplana belgica Houben, Proesmans & Artois, 2022
|
publication ID |
https://doi.org/10.5852/ejt.2022.798.1671 |
|
publication LSID |
lsid:zoobank.org:pub:F136E044-62C8-4FB3-8160-7DAE663D9600 |
|
DOI |
https://doi.org/10.5281/zenodo.6328786 |
|
persistent identifier |
https://treatment.plazi.org/id/1F08C372-CBAE-45E6-BE50-144752F3D44B |
|
taxon LSID |
lsid:zoobank.org:act:1F08C372-CBAE-45E6-BE50-144752F3D44B |
|
treatment provided by |
Felipe |
|
scientific name |
Bryoplana belgica Houben, Proesmans & Artois |
| status |
sp. nov. |
Bryoplana belgica Houben, Proesmans & Artois sp. nov.
urn:lsid:zoobank.org:act:
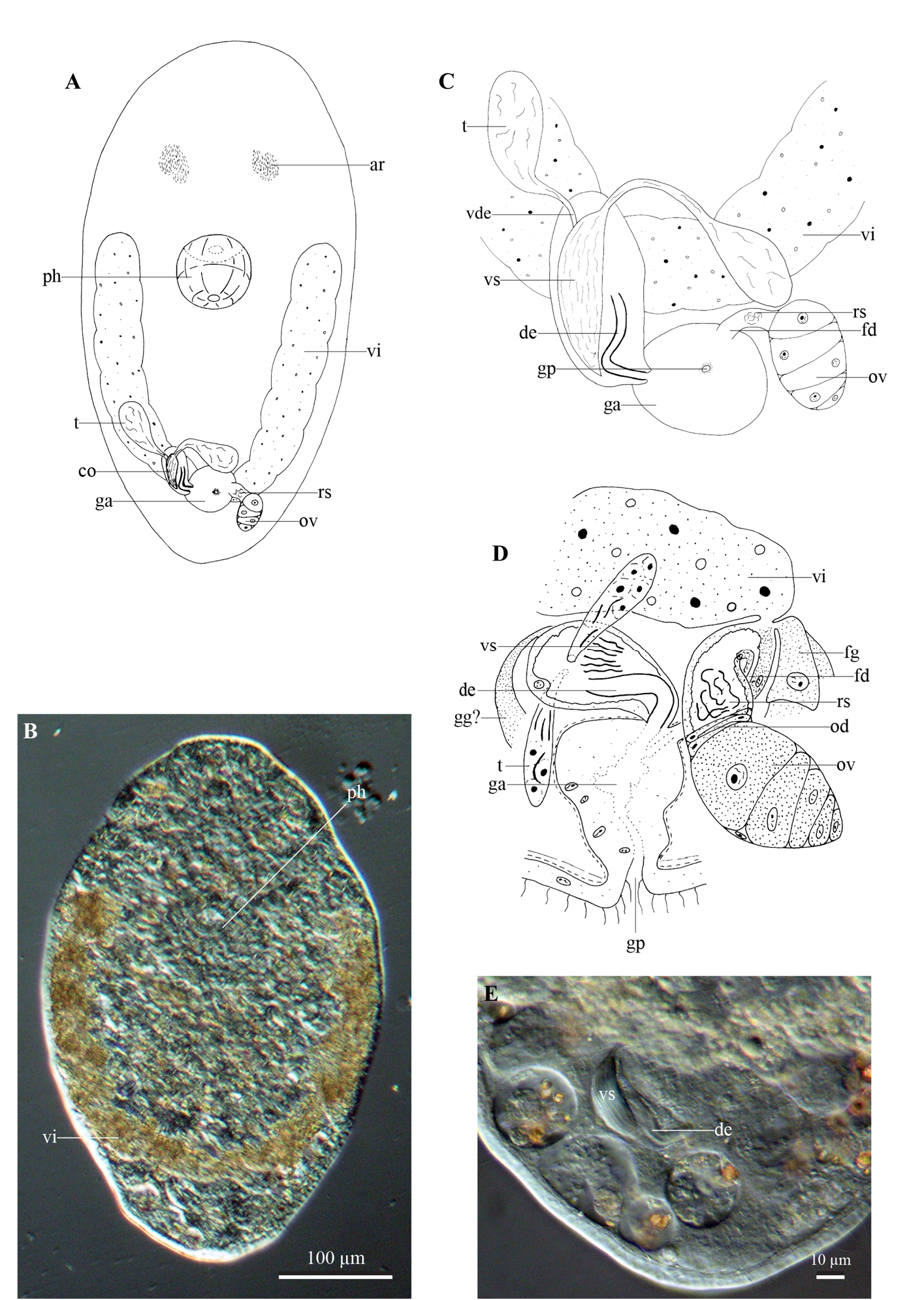
Fig. 5 View Fig
Diagnosis
Species of Bryoplana with the ciliation reduced or completely lacking dorsally. Testes elongated, in the posterior body part, lateral to the copulatory organ. Copulatory organ with bent, sclerotised ejaculatory duct. Most proximal part of female duct forming a seminal receptacle.
Etymology
The epithet refers to the fact that the species was found in Belgium.
Material examined
Holotype BELGIUM • 1 spec., studied alive and serially sectioned; Koksijde , Oostduinkerke, nature reserve ‘De Zeebermduinen’ ; 51°08′22″ N, 02°41′26″ E; 30 Jul. 2011; A.M. Houben and W. Proesmans leg.; dry moss growing on dunes; KV.687 ; FMNH. GoogleMaps
Description
The specimen is about 0.8 mm long. Both body ends are rounded ( Fig. 5A–B View Fig ). Free swimming specimens show a small tail. Adenal rhabdite glands ( Fig. 5A View Fig : ar) are situated in two groups at ±25% of the body. Dermal rhabdites and protonephridiopores were not observed. The epidermis on the ventral body side is 4.5 µm high and entirely covered with locomotory cilia, while on the dorsal body side it is 3.5 µm high and ciliation is strongly reduced, even almost completely lacking. A slightly forward-slanted rosulate pharynx ( Fig. 5A–B View Fig : ph) is located just rostral to the centre of the body.
The gonopore ( Fig. 5C–D View Fig : gp) is situated at ±80% of the body and connected to a genital atrium ( Fig. 5A, C–D View Fig : ga), which is lined with a high, nucleated epithelium and surrounded by muscles, the orientation of which could not be observed with certainty.
The elongated testes ( Fig. 5A, C–D View Fig : t) lie at ±70% of the body and ventrally to the paired vitellaria ( Fig. 5A–D View Fig : vi). They gradually taper into the broad vasa deferentia ( Fig. 5C View Fig : vde), which laterally enter the copulatory organ ( Fig. 5A View Fig : co). Circular muscles surround the 34 µm long copulatory organ, which includes an intracapsular seminal vesicle ( Fig. 5C–E View Fig : vs) and a 22 µm long, strongly sclerotised ejaculatory duct ( Fig. 5C–E View Fig : de). This ejaculatory duct is more or less straight at its proximal end and bends strongly (±100°) at the distal end. Coarse-grained eosinophilic glands are situated ventrally to the copulatory organ. Although the entrance into the copulatory organ could not be seen, these glands probably represent the prostate glands ( Fig. 5D View Fig : gg?).
The female duct ( Fig. 5C–D View Fig : fd) is relatively long and lined with a nuclear epithelium. Proximally, it widens into a seminal receptacle ( Fig. 5A, C–D View Fig : rs), which receives the short oviduct ( Fig. 5D View Fig : od). The vitelloduct and female glands ( Fig. 5D View Fig : fg) open into this female duct at the place where it connects to the seminal receptacle.
Discussion
The new species can readily be placed within ‘Typhloplanidae’ because it possesses all diagnostic features: a pharynx rosulatus, a single ovary, paired testes, and a single genital opening. Furthermore, the ventral position of the testes relative to the vitellaria indicates this species should be placed in ‘Protoplanellinae’, ‘Rhynchomesostominae’ Bresslau, 1933 or ‘Typhloplaninae’ Graff, 1905. However, the species’ general habitus and internal organisation differ markedly from the situation in the latter two subtaxa, and we therefore designate this species to ‘Protoplanellinae’.
Most representatives of ‘Protoplanellinae’ have the pharynx situated in the midbody or posterior body half. Only select species have a pharynx in the anterior half of the body, these belong to Achrochordonoposthia Reisinger, 1924; Bockia Reisinger, 1924; Bryoplana ; Microcalyptorhynchus Kepner & Ruebush, 1935, Prorhynchella Ruebush, 1939; and Protopharyngiellona Schwank, 1980. Some of these genera show very typical features: presence of a proboscis ( Microcalyptorhynchus), presence of ciliated pits in the anterior body half ( Prorhynchella), or the fact that the proboscis is of the doliiformis-type ( Bockia). Lack of these features in our species indicates that it cannot be allocated to any of these genera. One of the most eye-catching features of B. belgica Houben, Proesmans & Artois sp. nov. is the lack of a bursa copulatrix, which in the remaining taxa is only the case for Bryoplana . We hence allocate our specimens to the latter genus, to which it indeed shows much resemblance.
Bryoplana belgica Houben, Proesmans & Artois sp. nov. differs from Bryoplana xerophila by the presence of a sclerotised, curved ejaculatory duct, the presence of a seminal receptacle in the female system, and the fact that the dorsal body ciliation is very much reduced, or even lacking. The latter feature was also mentioned by Kolasa (1977) for Ventrociliella romanae, which he suggested to be an adaptation to limnoterrestrial habitats. However, in all other typical limnoterrestrial taxa, even in B. xerophila , the dorsal ciliation is present. Both species of Bryoplana were recovered from dried out moss – B. xerophila in the USA and B. belgica Houben, Proesmans & Artois sp. nov. in Western Europe – but indeed differ markedly in the extent of the dorsal body ciliation, undermining Kolasa’s (1977) hypothesis. The presence/absence of a seminal receptacle in the female system as a distinguishing character should be used with care at this moment. Indeed, the seminal receptacle in B. belgica Houben, Proesmans & Artois sp. nov. is simply a swelling of the female duct, and not a separate organ. As such, the absence of this structure in B. xerophila may be due to the specimens of Van Steenkiste et al. (2010) not having mated yet. However, as Van Steenkiste et al. (2010) investigated several live specimens and twelve serial sections, this seems unlikely. Regardless, the differences in body ciliation and construction of the copulatory organ proper clearly distinguish both species.
| FMNH |
Field Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Order |
|
|
Family |
|
|
Genus |