
Chorizogynopora italica Kolasa, 1981
|
publication ID |
https://doi.org/ 10.5852/ejt.2022.798.1671 |
|
publication LSID |
lsid:zoobank.org:pub:F136E044-62C8-4FB3-8160-7DAE663D9600 |
|
DOI |
https://doi.org/10.5281/zenodo.6328788 |
|
persistent identifier |
https://treatment.plazi.org/id/038A87DA-A777-FF8A-0413-FEC0FAF30EDD |
|
treatment provided by |
Felipe |
|
scientific name |
Chorizogynopora italica Kolasa, 1981 |
| status |
|
Chorizogynopora italica Kolasa, 1981 View in CoL
Fig. 6 View Fig
Material examined
GERMANY • 1 spec., studied alive; Schlitz-Breitenbach; 50°39′31″ N, 09°37′46″ E; 9 Aug. 2011; A.M. Houben and W. Proesmans leg.; mossy rocks GoogleMaps • 2 specs, studied alive; Wasserküppe; 50°29′27″ N, 09°56′52″ E; 10 Aug. 2011; A.M. Houben and W. Proesmans leg.; moss (Fontinalis sp.) near the source of the river Fulda GoogleMaps • 2 specs, studied alive and serially sectioned; same collection data as for preceding; 50°29′21″ N, 09°56′57″ E; XIV.2.39–XIV.2.40; HU GoogleMaps • 4 specs, studied alive, one of which serially sectioned; Graswiesenbach, Vogelsberg ; dark moss in fast-flowing water, on rocks in and just above the water; 50°32′08″ N, 09°12′00″ E; 10 Aug. 2011; A.M. Houben and W. Proesmans leg.; XIV.2.41; HU. GoogleMaps
Description and discussion
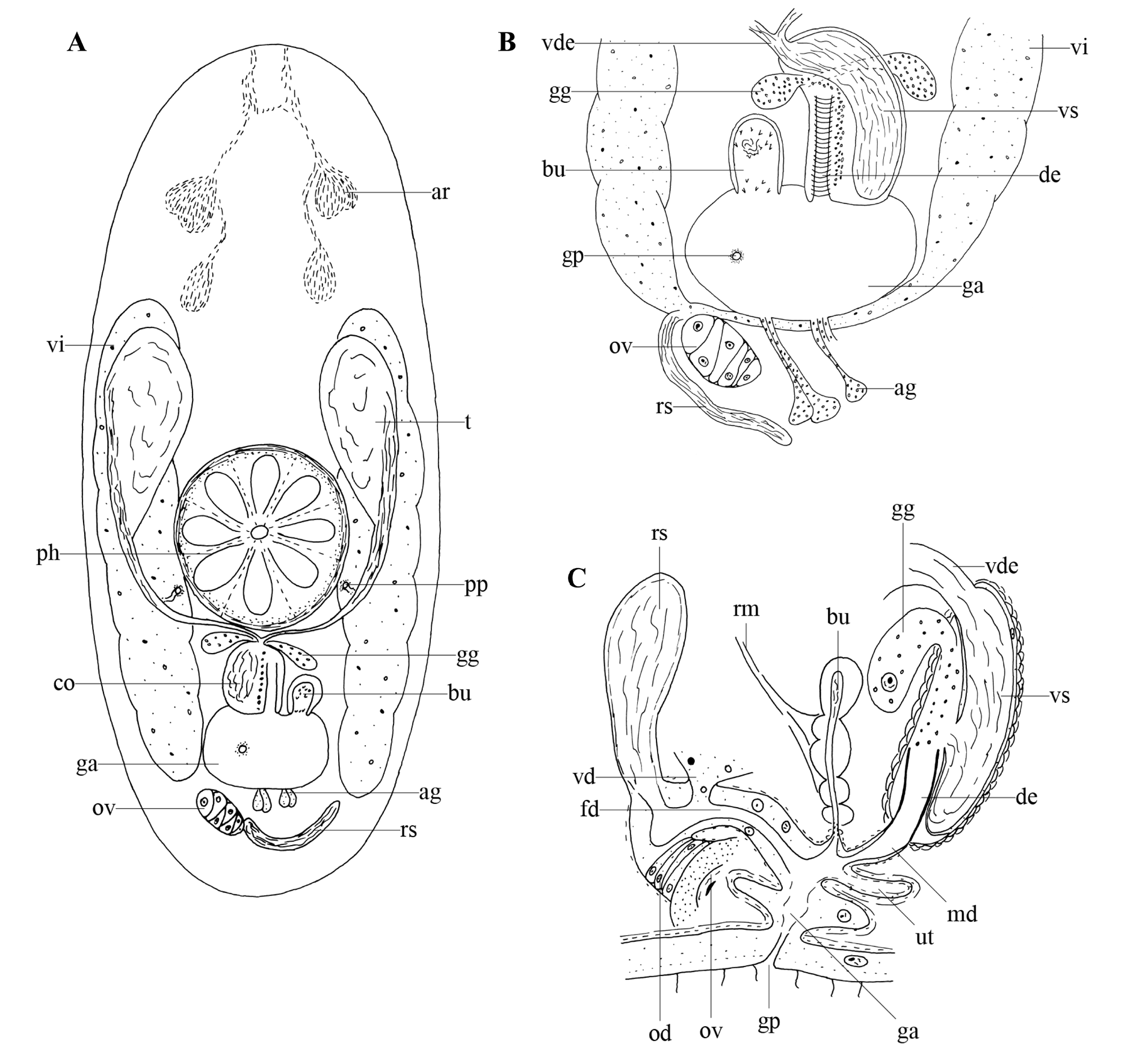
Habitus and internal organisation indicate that these specimens belong to Chorizogynopora italica. Although this species was properly described and illustrated by Kolasa (1981b), there are some small differences between our specimens and the original description. Therefore, we here provide a reconstruction of the genital system of our specimens.
The examined specimens are 0.7–0.8 mm long and very translucent. The body shape is blunt to rounded anteriorly and usually round with sometimes a small tail posteriorly ( Fig. 6A View Fig ). Adenal rhabdite glands ( Fig. 6A View Fig : ar) are situated at 25% of the body and give rise to two tracts that extend to the anterior body tip, partly anastomosing at their distal end. Dermal rhabdites were not observed. Paired protonephridiopores lie posterior to the rosulate pharynx ( Fig. 6A View Fig : ph), which lies just behind the centre of the body.
The gonopore ( Fig. 6A–C View Fig : gp) is situated at ±80% of the body and connected to a genital atrium ( Fig. 6A–C View Fig : ga). The latter is surrounded by an inner circular and outer longitudinal muscle layer.
The paired, large testes ( Fig. 6A View Fig : t) lie just anterior to the pharynx and ventral to the paired vitellaria ( Fig. 6A–B View Fig : vi). Both broad vasa deferentia ( Fig. 6B–C View Fig : vde) narrow gradually and fuse just before entering the copulatory bulbus. Two layers of spiral muscles surround the 52 µm long, oval-shaped copulatory organ ( Fig. 6A View Fig : co), which comprises an intracapsular seminal vesicle ( Fig. 6B–C View Fig : vs) and a slightly sclerotised ejaculatory duct ( Fig. 6B–C View Fig : de). The duct is surrounded by circular muscles (see Fig. 6B View Fig ). Large, coarse-grained, extracapsular eosinophilic glands ( Fig. 6A–C View Fig : gg) enter the copulatory organ at the proximal end. A short male duct ( Fig. 6C View Fig : md) connects the copulatory organ to the genital atrium. A small muscular bursa ( Fig. 6A–C View Fig : bu) containing sperm and prostate secretion is directly connected to the genital atrium. Its shape varies greatly between individuals. In some specimens, it is very narrow and elongate, while in others it is more egg-shaped. In some live specimens, groups of small spines were observed at both the proximal and distal end of the bursa, with the larger ones found at the distal end. Also associated with the bursa are some strong retractor muscles ( Fig. 6C View Fig : rm), which are anchored to the dorsal epidermis.
The vitellaria ( Fig. 6A–B View Fig : vi) extend at both sides of the body, from just behind the rhabdite glands to the level of the gonopore. The female duct ( Fig. 6C View Fig : fd) is relatively broad, surrounded by muscles and lined with a high, nuclear epithelium. It receives the oviduct ( Fig. 6C View Fig : od), a long seminal receptacle ( Fig. 6A–C View Fig : rs), and the common vitelloduct ( Fig. 6C View Fig : vd) proximally. A small muscular evagination of the atrium, which might serve as a uterus ( Fig. 6C View Fig : ut), is present.
The testes of our specimens are relatively larger, and the uterus relatively smaller than those described by Kolasa (1981b). Furthermore, Kolasa (1981b) observed tail glands, which we did not observe in our specimens. Despite these small differences the specimens studied are identified as C. italica due to their typical body shape, the presence of an elongated seminal receptacle, and the detailed structure of both the copulatory organ and the bursa.
Kolasa (1981b) described substantial differences between C. italica and C. paradoxa Reisinger, 1924, which is the only other species of Chorizogynopora Reisinger, 1924. Nevertheless, he distinctly mentioned these differences are probably not significant at the genus level. For a detailed discussion, the reader is referred to Kolasa (1981b).
Remarks
Specimens with one egg were observed. It was not possible to ascertain whether it was carried in the atrium or the uterus. Nematodes were observed in the gut of specimens.
Previously known distribution
Tuscany, Italy, in heavily moistened mosses growing in the stream Fosso Contesora ( Kolasa 1981b).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Order |
|
|
Family |
|
|
Genus |