
Mentocrex beankaensis, Goodman, Steven M., Raherilalao, Marie Jeanne & Block, Nicholas L., 2011
|
publication ID |
https://doi.org/10.5281/zenodo.206909 |
|
DOI |
https://doi.org/10.5281/zenodo.6188218 |
|
persistent identifier |
https://treatment.plazi.org/id/038C87A0-FFA5-6C5C-FF2B-1BDC3E5F5A3D |
|
treatment provided by |
Plazi |
|
scientific name |
Mentocrex beankaensis |
| status |
sp. nov. |
Mentocrex beankaensis , sp. nov.
Tsingy Wood Rail (English)
Râle des Tsingy (French)
Tsikozanalan’i Tsingy ( Malagasy)
Holotype. Département de Biologie Animale, Université d’Antananarivo ( UADBA 48179). Adult male collected by S. M. Goodman and M. J. Raherilalao at Madagascar: Province de Mahajanga, District de Maintirano, Forêt de Beanka, 4.9 km S. Ambinda, 18°3.7’S, 44°31.5’E, 320 m, on 31 October 2009. The specimen bears the field number MJR 866 and was used in both the morphological and molecular analyses.
Paratype. The Field Museum of Natural History ( FMNH 431145). Female downy young collected by S. M. Goodman and D. W. Willard at Madagascar: Province de Mahajanga, Parc National de Bemaraha, S bank Manambolo River, 3.5 km NE Bekopaka, 19°08.4’S, 44°49.7’E, 100 m, on 4 December 2001. The specimen bears the field number DW 5363 and was used in both the morphological and molecular analyses.
Etymology. The specific name is derived from the holotypical locality, Forêt de Beanka. The roots of this compound word are from the Malagasy “hanka”, which is the vernacular name of the Madagascar Long-eared Owl ( Asio madagascariensis ), and “be”, which refers to many. Hence, the word means the place where this owl is common.
Diagnosis. Mentocrex beankaensis , occurring in the central west of Madagascar, is distinctly larger than the two subspecies of M. kioloides , found in the east and northwest, in wing, tail and tarsus measurements. There are also distinct plumage coloration differences, most notably that in M. beankaensis the Cinnamon-Rufous (40) throat and moustachial stripe abut the lower portion of the eye, and the lores and forehead are mixed Smoke Gray (45) and Dark Grayish Brown (20).
Description of holotype. Lores, forehead, crown and upper nape a mixture of Smoke Gray (45) and Dark Grayish Brown (20) feathers, giving a mottled appearance ( Figs. 2 View FIGURE 2 , 3 View FIGURE 3 a). Ear-coverts and moustachial region dark Cinnamon-Rufous (40). Chin and throat off-white and bordered proximally by narrow Blackish Neutral Gray (82) line demarcating the division of the proximal limit of the throat, which terminates in a pointed shape. Breast and upper flanks dark Cinnamon-Rufous (40). Central portion of belly dark Clay Color (26) merging to tri-colored lower flanks composed of individual feathers barred with Dusky Brown (19), Raw Umber (23) and light Buff (24). Undertail coverts barred Raw Umber (23) and mixed with Tawny (38) and Cinnamon-Rufous (40). Thighs and tarsal feathering a mixture of feathers barred with Dusky Brown (19) and off-white. Axillaries, underwing coverts and flight feathers broadly barred back and white. Lower nape dark Clay Color (26), grading into a color approaching a light Raw Umber (23) on the mantle. Lower back, scapulars and wing coverts Amber (36) of slightly different intensities. Rump, uppertail coverts, uppertail and undertail Ferruginous (41).
Measurements of holotype. Wing (flattened) 142 mm, tail length 65 mm, tarsus length 46.1 mm, exposed culmen 27.1 mm, bill from anterior nostril 17.0 mm, bill width at anterior nostril 4.6 mm, weight 180 g ( Table 1).
Other details associated with the holotype. The following details are as noted on the specimen label of the holotype (UADBA 48179), which is an adult male (our translation from the French): iris—reddish-chestnut; man-dible—distal half black, proximal half whitish-gray; maxilla—whitish-gray; tarsus—black; stomach—empty; hab-itat—obtained in slightly disturbed dry deciduous valley forest with trees reaching 20 m in height; skull—fully ossified; gonads (testes)—left 4 x 3 mm, right 3 x 2 mm.
Description of paratype. The following details were noted on the soft part coloration of the paratype (FMNH 431145), which is a downy young female: iris—dark brown, mandible and maxilla—black with pale tip, tarsusbrownish black. The plumage coloration of this animal is seemingly identical to M. kioloides based on the illustration in Olson (1973, frontispiece).
Comparisons. There are notable differences in the head coloration between Mentocrex beankaensis and the two forms of M. kioloides ( Fig. 3 View FIGURE 3 a–c). In M. k. kioloides and M. k. berliozi the dark Cinnamon-Rufous (40) associated with the outer portion of the throat and moustachial stripe ends below the eye, while in the holotype of M. beankaensis this coloration abuts the lower portion of the eye. There are a few exceptions amongst the specimens of M. k. berliozi we have examined, with the dark Cinnamon-Rufous (40) nearly reaching the lower limit of the eye, but a small lores is clearly visible (e.g., AMNH 410370, 410376). In M. k. kioloides and M. k. berliozi , the lores and forehead are distinct, tending towards Olive-Gray (42) in the former and Smoke Gray (44) in the latter, as compared to the mixture of Smoke Gray (45) and Dark Grayish Brown (20) in M. beankaensis . In some individuals of M. k. kioloides , the forehead is slightly mottled with darker gray feathers (e.g., UADBA 48178, AMNH 410358) and the crown and upper nape are a dark Olive-Brown (28) as compared to a Fawn Color (25) or Clay Color (26) in M. k. berliozi or the mixture of Smoke Gray (45) and Dark Grayish Brown (20) in M. beankaensis . The ear coverts in M. k. kioloides and M. k. berliozi form a continuation of the same gray color passing posteriorly from the lores and below the eye, while in M. beankaensis this area is a continuation of the dark Cinnamon-Rufous (40) of the throat. The white portion of the throat in M. k. kioloides and M. k. berliozi is notably more extensive than in M. beankaensis , and in all three forms the proximal portion is delimited with a variable dark border that may be associated with how a given specimen was prepared.
In all three forms breast and upper body flanks are Cinnamon-Rufous (40), but based on the material available to us this is darker and more saturated in M. beankaensis , moderate in M. k. kioloides and lighter in M. k. berliozi . In this latter form the central portion of the belly is distinctly lighter than M. beankaensis , approaching a light Clay Color (26) with Buff Yellow (53) feather tips, and in the nominate form numerous individuals show light Buff (24) or whitish-cream feather tips. Undertail coverts in M. k. kioloides and M. k. berliozi are similar to M. beankaensis , but in the nominate form the Cinnamon-Rufous (40) tends to be more saturated.
The thigh and tarsal feathers in M. k. kioloides and M. k. berliozi are as in M. beankaensis , being dominated by Dusky Brown (19), but with a greater percentage of white barring. Axillaries, underwing coverts and flight feathers in M. k. kioloides and M. k. berliozi are similar to M. beankaensis , being broadly barred back and white. The lower nape in M. k. kioloides is a lighter Clay Color (26) than in M. beankaensis , while in M. k. berliozi it is dark Buff- Yellow (53) to light pinkish Clay Color (26). The mantle in M. k. berliozi is distinctly lighter, approaching dark Buff (24) or Clay Color (26), as compared to the other two forms in which it is light Raw Umber (23). Lower back, scapulars and upperwing coverts of M. k. berliozi approximate light Clay Color (26), as compared to the Amber (36) of the other two subspecies. Rump, uppertail coverts and rectrices of M. k. berliozi are light Ferruginous (41), as compared to a more saturated coloration in M. k. kioloides and M. beankaensis . On the basis of plumage coloration, no noticeable differences were found between a downy young of M. k. kioloides (AMNH 410365) and that of M. beankaensis (FMNH 431145).
Certain external measurements of the holotype of M. beankaensis (UADBA 48179) indicate that it is notably larger than M. kioloides ( Table 1). For the wing, tail and tarsus measurements, there is no overlap between these two species. In the case of the three different bill measurements, M. beankaensis falls within the range of M. kioloides . Given that only a single adult specimen is available of M. beankaensis , and that it is notably large, no statistical analyses have been conducted, as these would not be more meaningful than simple examination of the different measurement values.
Very few of the museum specimens available have detailed information on soft part coloration. The holotype of M. beankaensis (UADBA 48179) has a bicolored mandible, with the proximal half being blackish, the distal grayish-white and the leading edge grayish-white; the maxilla is grayish-white; and the tarsi black. In contrast, a specimen (FMNH 345622) referable to M. k. kioloides has the mandible and maxilla bluish-gray proximally, merging to a dull yellow tip, and the tarsi are black. Notes on labels of other specimens referred to the nominate form include: bill—blackish above, whitish below, legs—brownish-black (female, MNHN 1932.497); bill—black upper and bluish-white lower (female, MNHN 1932.496); bill—grayish base upper, blackish lower, legs—black (female, BMNH 1931.1008); bill—blue (male, BMNH 91.8.1.81); bill—pearl grey, legs—pearl grey (male, BMNH 1969.45.22). Whether these morphological differences are seasonal, sexual or individual will require further data from the field.
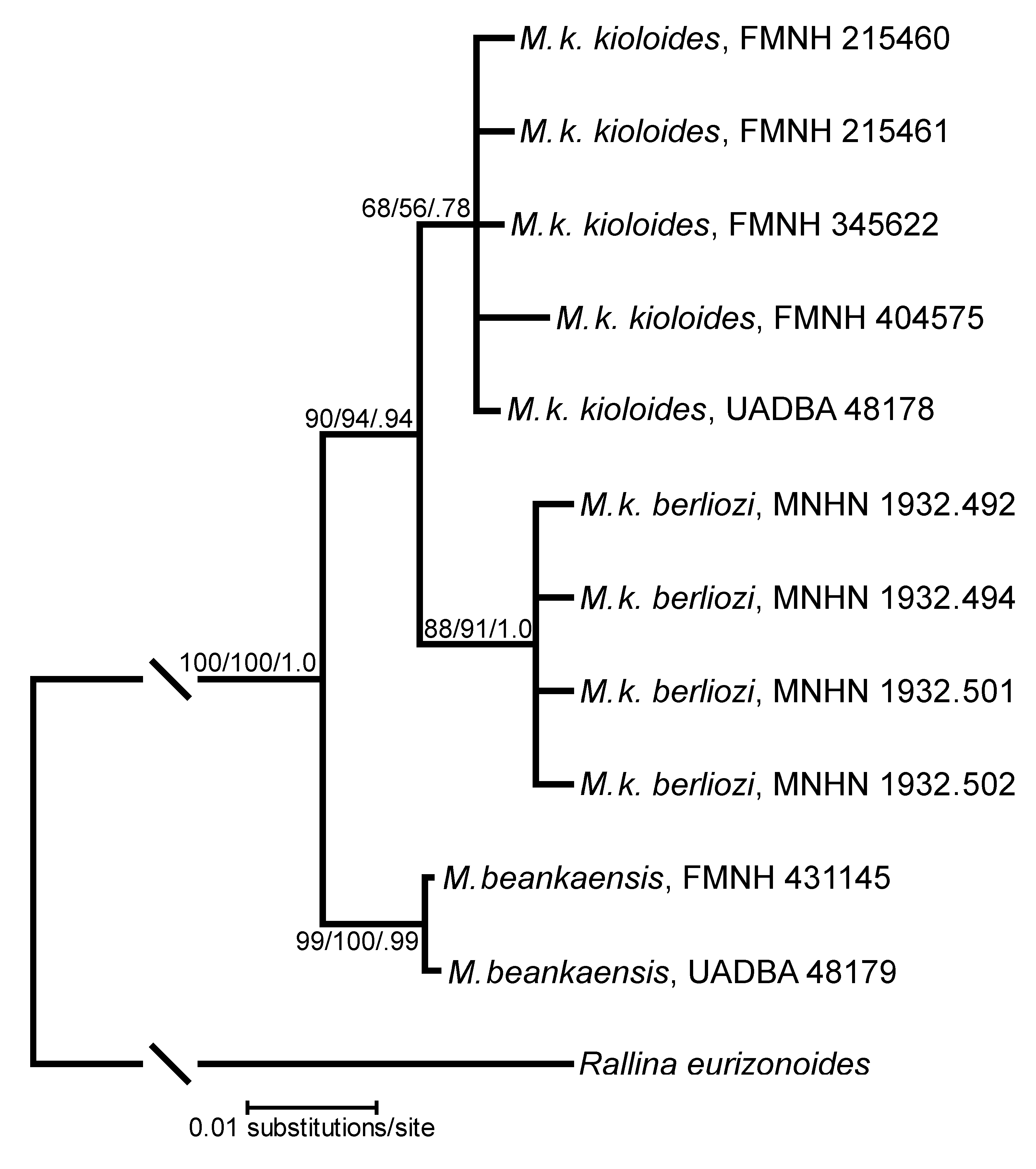
Although we examined a large percentage of the specimens of Mentocrex available in the world’s museums, the sample sizes are insufficient from most localities or too widely dispersed to properly assess patterns of geographical variation in size and coloration. Some conclusions can be presented here. Within specimens of M. kioloides from eastern Madagascar little morphological variation was found and these animals are referable to the nominate subspecies ( Table 1). The exceptions are three small adult specimens from extreme southeastern Madagascar, specifically the localities of Bemangidy (FMNH 215460, 215461) and Eminiminy (AMNH 703565). These birds have notably small bills, short wings, and more richly saturated Cinnamon-Rufous undersides. In contrast, those specimens from the northwestern portion of the island, generally considered the form M. k. berliozi , have different plumage patterns, particularly the head, upper back and breast and tend to be slightly larger than M. k. kioloides ( Figure 3 View FIGURE 3 , Table 1). The molecular analysis presented herein supports the separation of these two forms ( Figure 4 View FIGURE 4 ), but the level of differentiation is small enough to retain them as subspecies.
Molecular genetics. Phylogenetic tree constructions from MP, ML and BI analyses of the full dataset yield the same topology ( Fig. 4 View FIGURE 4 ). The ND3-only dataset has an identical topology but with slightly lower nodal support values (not shown). The two samples of M. beankaensis form a well-supported clade sister to a clade comprising M. k. kioloides and M. k. berliozi . Those two subspecies are also reciprocally monophyletic, although the support for the two clades is not very strong. Additional sequence data may strengthen these nodes, but the lack of any samples from the far northern range of M. k. kioloides leaves the exact genetic relationship between these subspecies unclear. Combined with the morphological data, the molecular genetic results support the treatment of M. k. kioloides and M. k. berliozi as subspecies.
For the ND3-only dataset, the average uncorrected p -distance between M. beankaensis and the kioloides / berliozi clade is 0.015 ± 0.005 SE (0.013–0.016), while it is 0.019 ± 0.003 (0.019–0.020) for the full dataset. Assessed separately, cytB’s p -distance between the clades ranges from 0.020–0.022, while ATP6’s ranges from 0.018–0.021. These full-dataset levels would likely increase with the addition of M. k. berliozi samples, as they were the most distant at ND3. Although species-level molecular data for Rallidae are scarce, the limited available data show that these p -distances are similar to those between other sister species in the family (J.M. Maley, pers. comm.; Trewick 1997; Hebert et al. 2004; Tavares et al. 2010).
Eight sites vary among the 372 bp of ND3, with four of the sites exhibiting fixed differences that diagnose M. beankaensis (Table 2). Three of the eight mutations resulted in amino acid substitutions as well, and all three are among the four that diagnose M. beankaensis . Of the 15 variable sites among the 654 bp of ATP6, 11 are fixed differences (Table 2). Four of the mutations caused amino acid substitutions, with three among the fixed differences. Finally, with cytB’s 917 bp, 17 of 21 variable sites are fixed. Only three changes were nonsynonymous, with two being fixed.
Distribution. Currently, Mentocrex beankaensis is known from a limited area of lowland central-western Madagascar, specifically portions of the Bemaraha and Beanka Massifs. This species was found at elevations from 100 to 320 m in areas of limestone karst characterized by rock pinnacles known in Malagasy as tsingy, often in canyons or valleys closed or bordered by exposed rock, and with dry deciduous forest that is more mesic than nearby more open formations.
The nominate form of kioloides has a broad distribution across the eastern length of the island, being known from lowland forest habitats near sea level to montane forest up to around 2000 m ( Morris & Hawkins 1998). We are unaware of any verified reports from the central portion of the Central Highlands, although it is known from numerous sites along the eastern edge of the Central Highlands (e.g., Raherilalao et al. 2007). We were unable to locate the specimen collected and reported by Salvan (1970) 30 km north of Ankazobe at 1450 m, which would be in the general vicinity of the Réserve Spéciale d’Ambohitantely, but as suggested by Benson et al. (1976), based on measurements, it might be referable to Dryolimnas cuvieri .
Mentocrex k. berliozi is known from the Sambirano area of the northwest. All examined specimens referable to this form were obtained in late 1930 and early 1931 during the Mission Zoologique Franco-Anglo-Américaine ( Rand 1936). In 1988, an expedition to this region working the western slopes of the Manongarivo Massif, between 120 and 400 m, did not find Mentocrex ( Thompson & Buisson 1988) . In 1999, a biological inventory of the northern slopes of the massif found this species in the zone between 785 and 1240 m ( Raherilalao et al. 2002), and those birds were tentatively assigned to M. k. berliozi .
Nomenclatural considerations. The name griseofrons Gray 1846 is a junior synonym of kioloides ( Sharpe 1894) . The illustration associated with the description of griseofrons ( Gray 1846, pl. 161) clearly shows the reduced amount of Cinnamon-Rufous on the outer portion of the throat and moustachial stripe, diagnostic of the nominate form. Hence, the name griseofrons is not applicable to the central western population described herein as M. beankaensis .
Conservation status. Few details are available on the distribution of Mentocrex beankaensis . It has been observed in different portions of the Bemaraha Massif. The direct line distance between the southernmost known record in the Bemaraha and the site at Beanka is 125 km and the width of appropriate habitat across this zone does not exceed 5 km in most areas. Hence, based on current information this species has a very limited distribution. Further research is needed to define its known geographical range more clearly and to derive estimates of population density. Most of the Bemaraha Massif falls within the protected areas system of Madagascar and the terra typica of this new species is under the conservation management by the organization Biodiversity Conservation Madagascar (BCM).


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.