
Hongshanopterus lacustris, Wang, Xiaolin, Campos, Diogenes De Almeida, Zhou, Zhonghe & Kellner, Alexander W. A., 2008
|
publication ID |
https://doi.org/10.5281/zenodo.274343 |
|
DOI |
https://doi.org/10.5281/zenodo.6232917 |
|
persistent identifier |
https://treatment.plazi.org/id/038D0E22-FF88-FFEA-FF5C-FA8ABEB1651B |
|
treatment provided by |
Plazi |
|
scientific name |
Hongshanopterus lacustris |
| status |
sp. nov. |
Hongshanopterus lacustris sp. nov.
Etymology. The specific name comes from the Latin word lacustris , ‘lacustrine’, in reference to the depositional environment of some layers of the Jiufotang Formation.
Holotype. A skull, atlas, axis and three other cervical vertebrae embedded in a shale slab, housed at the Institute of Vertebrate Paleontology and Paleoanthropology of the Chinese Academy of Sciences, Beijing, China ( IVPP V14582 View Materials ; Figs. 1–11 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8 View FIGURE 9 View FIGURE 10 View FIGURE 11 ).
Type locality and horizon. Dapingfang, Chaoyang, western Liaoning, China. Jiufotang Formation, Aptian, Lower Cretaceous ( He et al. 2004).
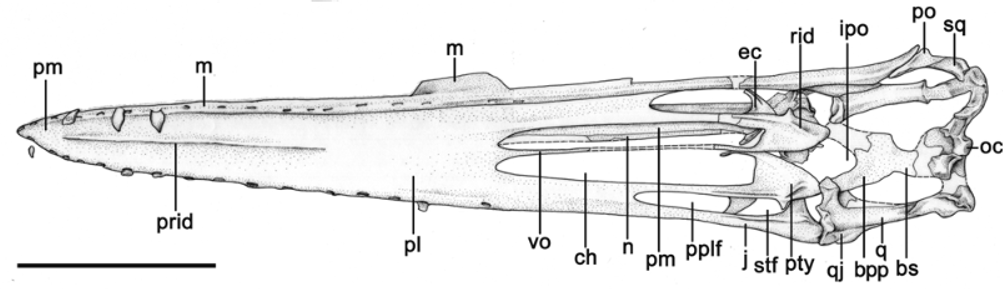
Diagnosis. An istiodactylid pterosaur distinguished from all other members of this clade by the following combination of characters: comparative large number of upper teeth (34–38) more than in any other istiodactylid); dentition present until a little over half of the skull length (pm-sq); presence of teeth with the crown distinctively directed posteriorly; pterygoid with a ventral anterolateral directed ridge ( Figs. 1–11 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8 View FIGURE 9 View FIGURE 10 View FIGURE 11 ).
Description. Generalities and taphonomy. The specimen, embedded in a beige colored shale slab, is composed of the skull and five isolated cervical vertebrae (including the atlas and axis) that due to their close association are regarded as belonging to the same animal ( Fig. 1 View FIGURE 1 ). The skull can only be seen in palatal view. It was compressed dorsoventrally during the fossilization process, causing a deformation from left to right, with some cranial bones displaced to the left side. Due to this displacement, the internal surface of some elements of the skull roof can be seen inside the choana, subtemporal and postpalatinal fenestrae of the left side ( Figs. 2 View FIGURE 2 , 3 View FIGURE 3 ). The atlas-axis complex together with two cervical vertebrae was displaced close to the right anterior tip of the skull. The atlas-axis overlies the posterior part of one cervical vertebra. A fifth cervical vertebra is found posterior to the occipital region of the skull and is not well preserved. The bone surface in most elements is broken in tiny pieces, reminding of the crackled shale of a broken egg, which is commonly observed in pterosaur specimens. This is particularly true for the anterior palatal region that contrasts with the comparatively better preserved bone surface of the posterior portion of the skull ( Figs. 4 View FIGURE 4 , 5 View FIGURE 5 ).
Based on the observations above, it is not clear if the carcass reached the bottom of the water column as one piece or if it was dismembered somewhere else. In any case, at least part of the neck and the skull of this pterosaur reached the bottom of the water column and remained exposed for some time, with soft tissue rotting away and bones being dismembered and displaced from their original anatomical position (e.g., atlas+ axis and other cervical vertebrae). The disposition of the bones and the fact that the cervical vertebrae were not carried away is suggestive of a low-energy environment.
Skull and cranial openings. The skull is long, a typical feature of pterosaurs and terminates in a pointed end, lacking any lateral expansion. The orbit and nasoantorbital fenestra are not visible due to crushing but the left upper and lower temporal fenestra are partially preserved ( Figs. 1–3 View FIGURE 1 View FIGURE 2 View FIGURE 3 ). The upper temporal fenestra is the smallest and judging from the preserved portion is oriented dorsally with a rounded outline. The lower temporal fenestra, although taphonomically deformed, is quite elongated, more than in the Anhangueridae (e.g., Kellner & Tomida 2000) and Pteranodon ( Bennett 2001) . The choanae are elongated, reaching approximately 27.8% of the skull length measured from the tip of the premaxillae to the posterior margin of the squamosal (pm-sq; see Table 1). They are the largest openings in the palate, with the anterior and part of the lateral margin formed by the palatine and the lateroposterior margin formed by the pterygoid. The choanae are separated from each other by thin vomers. The postpalatinal fenestra is much smaller and shows a rather sub-triangular outline. It is perforated by a developed lateral process of the pterygoid. The interpterygoid opening is well developed.
Length from the tip of premaxilla to the squamosal—243.5 Length from quadrate to premaxilla—206.7
Length choana (right side)—67.7
Length premaxilla to the anterior margin of the choana (right side)—119.2 Length of postpalatinal fenestra—26.5
Length of subtemporal fenestra—21.2
Width over quadrates (over the ventral part of the lateral articulation surfaces)—39.8 Premaxilla and maxilla. There is no suture visible between the premaxilla and maxilla and the exact number of premaxillary teeth cannot be established. The anterior portion of the upper jaw is pointed and lacks any lateral expansion ( Fig. 4 View FIGURE 4 ). The tooth bearing margin, better preserved on the left side, is formed by thick bone raised in respect to other palatal elements. The limit of maxilla with the palatine is marked by a deep sulcus, but the posterior boundaries of this bone cannot be established. Based on the presence of a developed sulcus on the medial surface of the right jugal, it is possible that the maxilla sends a posterior ramus that takes part in the lateral margin of the subtemporal fenestra. On the left side there are at least 17 alveoli and possibly two more, indicating the presence of 17 to 19 teeth on each side of the upper jaw (see dentition). The alveoli are elongated anteroposteriorly. They tend to be projected and show thickened borders. The alveoli are overall evenly spaced and the spacing in most cases is larger than the longest axis of the adjacent alveolus.
The ventral surface of two elongated bones can be observed inside the left choana ( Figs. 2 View FIGURE 2 , 3 View FIGURE 3 ). Based on their position, they are interpreted as the premaxilla and nasal from the right side. This portion of the premaxilla suggests that the nasoantorbital fenestra is well developed.
Nasal. A strip-like bone is observed inside the left choana and is interpreted as the right nasal ( Figs. 2 View FIGURE 2 , 3 View FIGURE 3 ). This bone contacts the premaxilla ventrolaterally but, despite being elongated, probably did not reach the most anterior portion of the nasoantorbital fenestra. The posterior portion of the nasal cannot be observed.
Squamosal. The squamosal is not well preserved ( Figs. 2 View FIGURE 2 , 3 View FIGURE 3 , 6 View FIGURE 6 ). If forms the lateral margin of the posterior end of the skull, the posterolateral margin of the upper temporal fenestra and the posterodorsal margin of the lower temporal fenestra. The suture between this bone and the jugal is marked by a line on the left side.
Jugal. The right jugal is the best preserved of the two and was displaced medially, inside the posterior end of the nasoantorbital fenestra ( Figs. 2 View FIGURE 2 , 3 View FIGURE 3 , 5 View FIGURE 5 , 7 View FIGURE 7 ). The lacrimal process has a broad base, and it is not clear if it thins upward and is posteriorly inclined in the same fashion as observed in other istiodactylids ( Wang et al. 2005; Andres & Ji 2006). The right jugal shows a well developed sulcus on the medial surface, possible to receive the jugal ramus of the maxilla. The jugal has a strong postorbital process, which can be seen on the left side. In this region, a small portion of the postorbital is observable.
Quadratojugal. The quadratojugal on the left side is better preserved than on the right ( Figs. 2 View FIGURE 2 , 3 View FIGURE 3 , 6 View FIGURE 6 ). It is a small bone that forms the ventral margin of the lower temporal opening, contacting the jugal dorsally and the quadrate ventrally. On both sides this bone is partially fused with the jugal and the quadrate and only a faint line indicates the sutures. In its ventral portion, the quadratojugal exhibits an odd ‘knob-like’ protuberance positioned close to the quadrate articulation ( Fig. 5 View FIGURE 5 ). The pterodactyloid Dsungaripterus (IVPP V64045 View Materials -9) also shows a protuberance in this region, but is more developed and placed more ventrally.
Quadrate. This bone is well preserved on both sides ( Figs. 2 View FIGURE 2 , 3 View FIGURE 3 , 5 View FIGURE 5 ). It is a long rod-like element that has a wide base for the mandibular articulation. It sends a thick medial process that articulates with the basipterygoid process of the basisphenoid and the pterygoid. On the right side, the quadrate shows a faint laminar bone, indicating the presence of a shallow embayment (e.g. quadrate flange), such as reported in anhanguerids ( Kellner & Tomida 2000) and other more derived pterodactyloids (e.g., Dsungaripterus , Pteranodon , Thalassodromeus ).
The articulation with the mandible has suffered some compression that has somewhat altered its shape (compare the right and left quadrates on Fig. 7 View FIGURE 7 ). The better preserved one is the right ( Fig. 5 View FIGURE 5 ). The articulation surface is formed by two concavities separated by a small ridge. The lateral concavity is larger and the medial one very shallow. This is similar to the condition reported in Anhanguera araripensis and Pteranodon (Wellnhofer 1985; Eaton 1910), which differ from the Chinese specimen by having the medial concavity more developed than the lateral one. This structure is commonly called the helical jaw joint ( Eaton 1910; Bennett 2001). Apparently the lower jaws of Liaoningopterus (originally regarded as the wing phalanges, Wang & Zhou 2003) also have a helical jaw joint, similar to Anhanguera ( Campos & Kellner 1985) .
Basisphenoid. The occipital region has suffered severely from compression and only some elements can be observed. The best preserved one is the basisphenoid ( Figs. 2 View FIGURE 2 , 3 View FIGURE 3 ) that forms a thin and fragile anteriorly directed bony plate. It bears short and broad lateroventrally processes (the basipterygoid processes) that contact the quadrate and the pterygoid. Although it is not possible to establish the exact length of this bone, it appears to be comparatively short.
Basioccipital and exoccipital. The basioccipital forms the rounded occipital condyle, and has strong depression below it ( Figs. 2 View FIGURE 2 , 3 View FIGURE 3 , 6 View FIGURE 6 ). The exoccipital is made of relatively thick bone and forms at least the lower half of the occipital region. An eye-shaped opening is observed on the right side, totally surrounded by this bone.
Palatine. The limits between the palatine and the surrounding bones cannot be determined easily in this specimen due to fusion ( Figs. 2 View FIGURE 2 , 3 View FIGURE 3 ). The suture with the maxilla is formed by a distinct groove that can be followed until the postpalatinal fenestra. No limits are discernible with the premaxillae or with the vomers. The limits with the pterygoid are also not very clear and in some parts are indicated by a faint groove. The palatine surrounds the anterior part of the choanae and also at least part of the postpalatinal fenestrae but appears not to contact the ectopterygoid. Anteriorly, a palatal ridge starts between the level of the 3rd and 4th alveoli ( Fig. 4 View FIGURE 4 ), gets more pronounced at the level of the 10th alveolus and decreases from there until fading away between the level of the 13th-14th alveoli.
Pterygoid. Both pterygoids are relatively well preserved ( Figs. 2 View FIGURE 2 , 3 View FIGURE 3 ). They form the posterior margin of the choanae, are unfused and meet at the midline forming two small anteriorly projected processes that are overlain by the vomers. The anteriorly tapering palatinal process forms the lateral margin for over the half length of the choana and part of the medial margin of the postpalatinal fenestra. Posteriorly this bone is connected with the quadrate and basisphenoid. The posterior processes diverge leaving a comparatively wide interpterygoid opening. Laterally this bone shows a well developed process that penetrates the subtemporal fenestra. A well developed anterolaterally directed ridge ( Fig. 5 View FIGURE 5 ), followed by a marked anteroposteriorly directed sulcus can be observed on both pterygoids ( Fig. 7 View FIGURE 7 ).
Ectopterygoid. The ectopterygoid is better preserved on the left side. This bone shows a marked sulcus and makes the boundary between the postpalatinal and subtemporal fenestrae ( Figs. 2 View FIGURE 2 , 3 View FIGURE 3 , 7 View FIGURE 7 ).
Vomer. The vomers are thin medial elements that posteriorly join the pterygoids and anteriorly the palatines. They divide the choanae, are thinnest at their middle point and gradually get wider posteriorly and anteriorly ( Figs. 2 View FIGURE 2 , 3 View FIGURE 3 ).
Teeth. Several alveoli show remains of teeth but only four are preserved enough to be described ( Fig. 4 View FIGURE 4 , 8 View FIGURE 8 ). Only part of the crown of the second tooth from the left side (likely from the premaxilla) is preserved indicating that this (and perhaps also the first) was short and more rounded compared to the others. The remaining three are all from the left side: two isolated (possible the fourth and seventh) and the eight, still in the alveolus ( Fig. 8 View FIGURE 8 ). All three teeth are strongly labiolingually compressed with pointed triangular crows. There is no marked constriction separating the crown from the root. The seventh has the crown curved posteriorly. In all these teeth the crown is rather smooth and lacks carinae. As far observation is possible, the roots are subequal in size with the crown. The dentition extends after the anterior margin of the choanae.
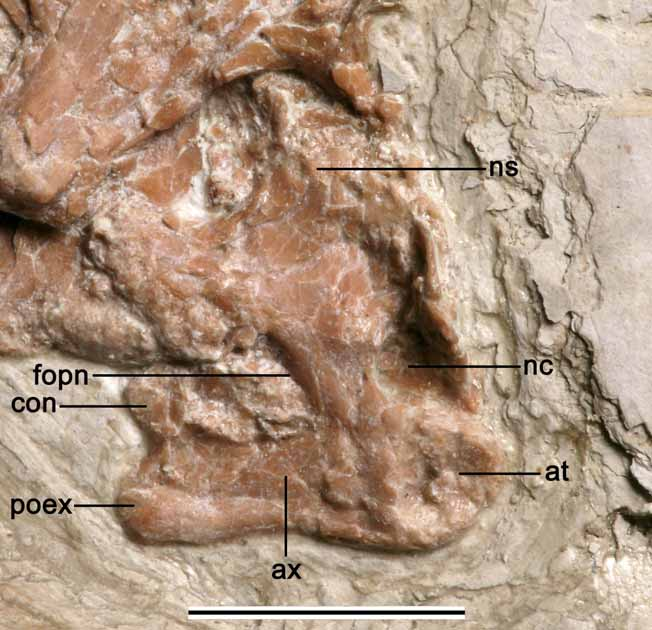
Cervical Vertebrae. In total, five cervical vertebrae were found ( Fig. 1 View FIGURE 1 ). The atlas and axis were displaced over the posterior end of an anterior cervical ( Fig. 9 View FIGURE 9 ). The atlas is apparently fused to the axis and the limits of those elements cannot be established ( Fig. 10 View FIGURE 10 ). The combined length of the centrum is about 11 mm. There are some broken bone parts anterior to the neural spine of the axis and partially displaced inside the neural canal; it is not clear if they belong to the axis or the atlas. The anterior surface of the atlas shows a well developed rounded concavity that receives the occipital condyle. A blunt knob-like projection is observed on the right anterolateral surface, lateral to the neural canal and might be a region where the neural spine of the atlas is fused to the axis.
The axis is a strong element that has a well developed neural spine whose dorsal surface is broken. The postzygapophysis is strong and directed lateroposteriorly. The postexapophyses are also strongly developed. A well developed pneumatic foramen pierces the lateral side of this bone.
The remaining cervical vertebrae show basically the same morphology but vary in size. The largest one is in contact with the axis ( Fig. 9 View FIGURE 9 ) and has its lateral right side exposed (length between prezygapophysis and postexapophysis ~ 27 mm). It shows a high spike-like neural spine. The lateral side of the centrum is pierced by an eye-shaped pneumatic foramen.
Another cervical vertebra is positioned between the previous ones and the skull and is exposed in ventral view ( Fig. 1 View FIGURE 1 ). It is smaller (length of the centrum = 21.5 mm), clearly procoelic and shows a well developed hypapophysis ventral to the cotyle ( Fig. 11 View FIGURE 11 ). Despite being dorsoventrally compressed, the lateral surfaces show evidence of a pneumatic foramen.
The last preserved cervical vertebra is located close to the left rear end of the skull ( Fig. 1 View FIGURE 1 ). It is exposed in dorsal view and is the smallest element of all (estimated length of the centrum = 14 mm). None show evience of cervical ribs.









| IVPP |
Institute of Vertebrate Paleontology and Paleoanthropology |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |