
Leucascus simplex Dendy, 1892
|
publication ID |
https://doi.org/10.11646/zootaxa.3619.3.3 |
|
publication LSID |
lsid:zoobank.org:pub:92C07D63-F2F5-4898-A7FE-4937F4D5A043 |
|
DOI |
https://doi.org/10.5281/zenodo.6153178 |
|
persistent identifier |
https://treatment.plazi.org/id/038E706D-EC16-7440-FF17-2C0CC66EFD4B |
|
treatment provided by |
Plazi |
|
scientific name |
Leucascus simplex Dendy, 1892 |
| status |
|
Leucascus simplex Dendy, 1892 View in CoL
( type species, by subsequent designation; Dendy & Row 1913)
Diagnosis: Cortical, choanosomal, and atrial skeletons composed mainly of triactines. Apical actine of the tetractines ornamented with spines. Choanocyte tubes predominantly smooth.
Synonymies: Leucascus simplex Dendy 1892: 77 ; Kirk 1897: 313; Dendy 1913: 9; Dendy & Row 1913: 731; Row & Hôzawa 1931: 742.
Type material: BMNH 1925.11.1.1729 ( syntype; Port Phillip Heads, Australia; R.N. 229; Dendy collection), BMNH 1925.11.1.1730 ( syntype; Port Jackson, Australia; R.N. 217; Dendy collection), BMNH 1925.11.1.1731 ( syntype; Port Phillip, Australia; R.N. 226; Dendy collection), BMNH 1925.11.1.34 ( syntype; Port Phillip Heads, Australia; R.N. 230; Dendy collection), NMV F42418 View Materials ( syntype; fragment of the specimen BMNH 1925.11.1.1730), and NMV F42419 View Materials ( syntype; fragment of the specimen BMNH 1925.11.1.1731).
Type locality: Port Phillip Heads, Australia.
Additional analysed material: BMNH 1925.11.1.1399a (Hamburg, S.W. Australian Expedition, 1905; Dendy collection).
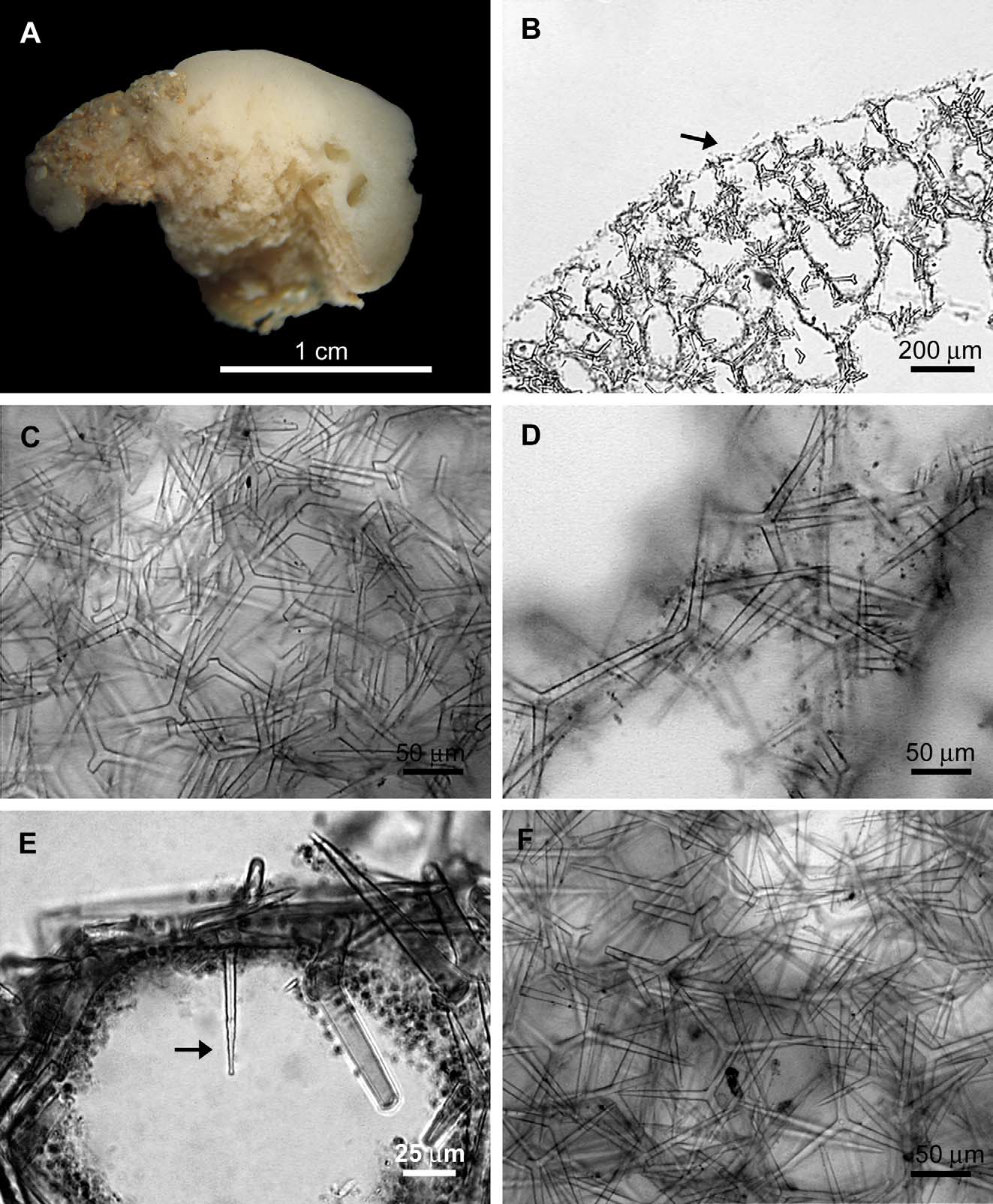
Description: The colour in vivo is unknown, but the preserved specimens are beige ( Figure 1 View FIGURE 1 A). The cormus is massive, formed by tightly anastomosed tubes. The consistency is firm but compressible. The cormus is covered by a thin membrane. In the preparations made by Dendy (BMNH 1925.11.1.1399a, and BMNH 1925.11.1.1731), the cortical membrane could be observed only in some regions ( Figure 1 View FIGURE 1 B). Nevertheless, it was easily observed in slides of the syntypes NMV F42418 View Materials and NMV F42419 View Materials . Oscula are simple apertures without any ornamentation and the atrium is reduced, surrounded by a membrane and predominantly smooth.
The skeletons of the cortex and tubes are formed mainly by triactines, but rare tetractines are also present ( Figures 1 View FIGURE 1 C, D). Consequently, tubes are predominantly smooth, but some of them are slightly hispid because of the apical actine of the rare tetractines ( Figure 1 View FIGURE 1 E). The atrial skeleton is similar to the cortical one, with abundant triactines and rare tetractines ( Figure 1 View FIGURE 1 F).
Spicules/ Specimens Actine Length (µm) Width (µm) N
Min. Mean SD Max. Min. Mean SD Max.
Triactine
BMNH 1925.11.1.34 83.2 102.9 6.7 114.4 11.7 13.3 0.9 15.9 30
BMNH 1925.11.1.1731 83.2 99.4 8.1 114.4 7.8 10.0 1.2 13.0 30
BMNH 25.11.1.1730 83.2 95.4 7.6 111.8 6.5 8.3 0.7 9.1 30
All specimens 83.2 99.2 3.1 114.4 6.5 10.5 2.1 15.9 –
Tetractine Basal
BMNH 1925.11.1.34 72.8 100.1 9.2 117.0 10.4 12.1 1.0 14.3 30
BMNH 1925.11.1.1731 85.8 98.8 5.9 109.2 7.8 9.0 0.8 10.4 30
BMNH 25.11.1.1730 83.2 96.8 7.3 111.8 7.8 8.3 0.7 10.4 30
All specimens 72.8 98.6 1.3 117.0 7.8 9.8 1.6 14.3 –
Apical
BMNH 1925.11.1.34 26.0 42.3 8.8 65.0 2.6 4.0 1.0 5.2 30
BMNH 1925.11.1.1731 43.7 56.9 9.6 85.0 3.7 4.8 0.5 6.1 30
BMNH 25.11.1.1730 21.9 33.8 9.1 43.7 2.4 3.3 0.9 4.9 12
All specimens 21.9 44.3 9.5 85.0 2.4 4.0 0.6 6.1 – Spicules ( Table 1):
( i) Triactines ( Figure 2 View FIGURE 2 A): Regular. Actines are conical, straight, with blunt tips;
( ii) Tetractines ( Figure 2 View FIGURE 2 A): Similar to the triactines, but with an apical actine, which is smaller and thinner than the basal actines and shows well developed and abundant spines ( Figures 2 View FIGURE 2 B).
Remarks: Differently of Dendy’s description (1892), the tetractines of Leucascus simplex do not present a vestigial apical actine. Considering the three syntypes measured in the present work, this actine has a mean value of 44.3 µm ( Table 1) and previous authors have already described long apical actines in this species. Kirk (1897), for example, found a value of 70.0 µm for specimens from New Zealand, and Dendy (1913) mentioned that the apical actine of specimens from Providence ( Seychelles) was well developed (but did not mention the values). Another important observation related to the apical actine of the tetractines was the presence of spines in all specimens studied. This character has previously not been observed in the genus Leucascus .
Leucascus neocaledonicus and L. clavatus are the species that are most similar to L. simplex . The only difference between L. simplex and L. neocaledonicus is the higher abundance of tetractines in L. neocaledonicus , which results in tubes that are more hispid. In relation to L. clavatus , it can be easily separated from L. simplex mainly by the presence of diactines in the former. Moreover, it has spicules with sharp tips and an atrial skeleton composed exclusively of triactines, while in L. simplex the tips are blunt and the atrial skeleton is formed mainly by triactines and scattered tetractines.
Distribution: Leucascus simplex is allegedly a cosmopolitan species. Nevertheless, up to date no species of Calcarea has proved to be truly cosmopolitan. In the present work we reject the previous records of this species in the Atlantic Ocean ( Brazil) (Borojevic & Peixinho 1976; Muricy et al. 1991). Contrary to L. simplex , the Brazilian specimens showed great abundance of tetractines in the atrial skeleton and microdiactines in several parts of the body. These specimens are being described here as Leucascus albus sp. nov. Therefore, up to date L. simplex is considered to be present in the Indian and Pacific Oceans: near Port Phillip Heads, Bass Straits, Watson’s Bay, Port Jackson (Dendy 1892); Shark Bay, Geraldton District, Fremantle District, Bunbury District (Row & Hôzawa 1931)— Australia; Seychelles (Dendy 1913), and New Zealand (Kirk 1897). Spalding et al. (2007) corresponding ecoregions are: Bassian, South Australian Gulfs, Manning-Hawkesbury, Shark Bay, Houtman, Leeuwin, Seychelles, and Central New Zealand.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |