
Ennominae Duponchel, 1845
|
publication ID |
https://doi.org/10.7717/peerj.7386 |
|
DOI |
https://doi.org/10.5281/zenodo.5767485 |
|
persistent identifier |
https://treatment.plazi.org/id/038F87AD-2B07-FFAB-2EC5-5947FB400E30 |
|
treatment provided by |
Carolina |
|
scientific name |
Ennominae Duponchel, 1845 |
| status |
|
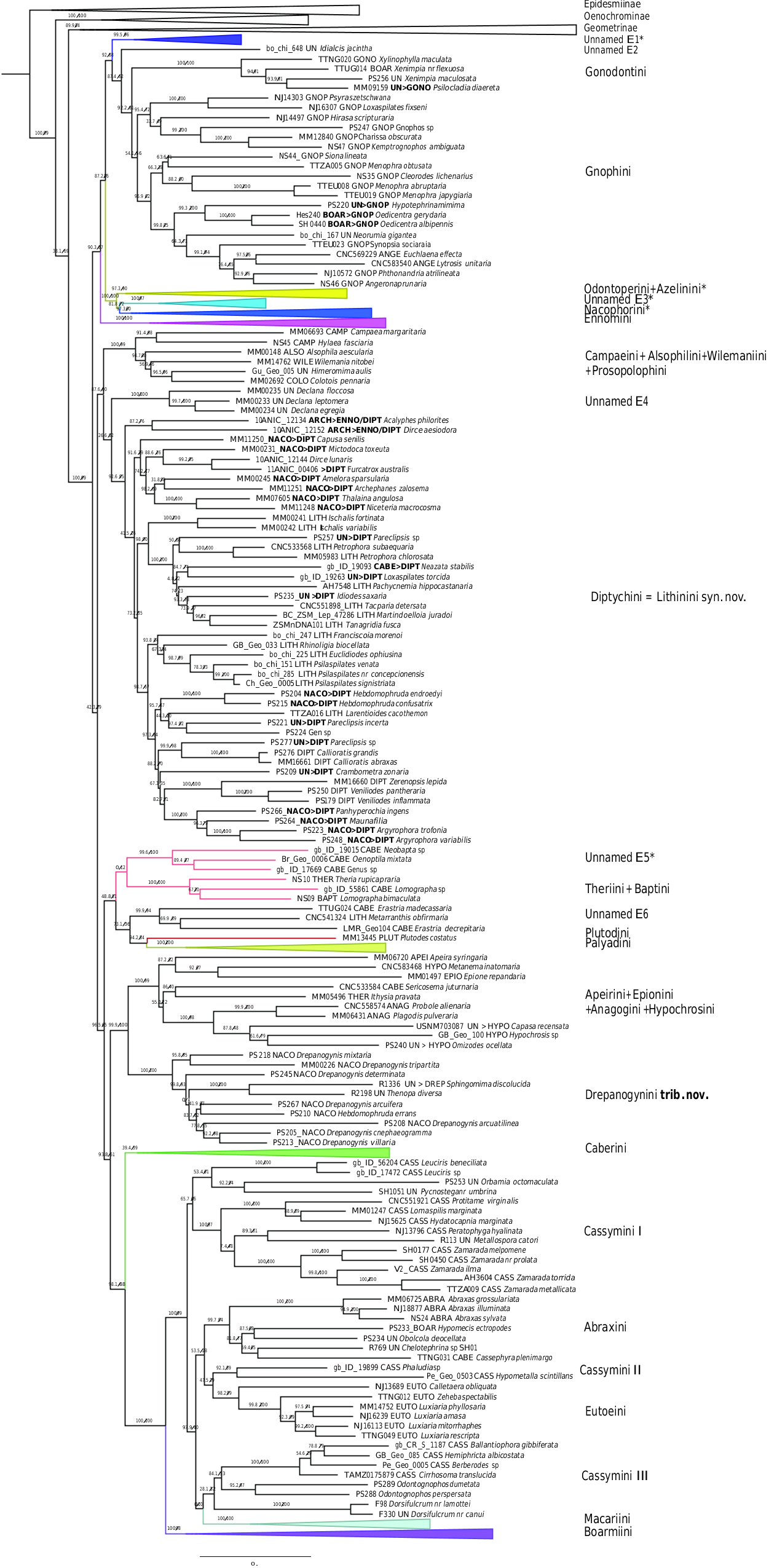
Ennominae are the most species-rich subfamily of geometrids. The loss of vein M2 on the hindwing is probably the best apomorphy ( Holloway, 1994), although vein M2 is present as tubular in a few ennomine taxa ( Staude, 2001; Skou & Sihvonen, 2015 ). Ennominae are a morphologically highly diverse subfamily, and attempts to find further synapomorphies shared by all major tribal groups have failed.
The number of tribes as well as phylogenetic relationships among tribes are still debated (see Skou & Sihvonen, 2015 for an overview). Moreover, the taxonomic knowledge of this subfamily in tropical regions is still poor. Holloway (1994) recognized 21 tribes, Beljaev (2006) 24 tribes, and Forum Herbulot (2007) 27 tribes. To date, four molecular studies have corroborated the monophyly of Ennominae ( Yamamoto & Sota, 2007; Wahlberg et al., 2010; Õunap et al., 2011, Sihvonen et al., 2011), with Young (2006) being the only exception who found Ennominae paraphyletic. Moreover, four large-scale taxonomic revisions (without a phylogenetic hypothesis) were published by Pitkin (2002) for the Neotropical region, Skou & Sihvonen (2015) , Müller et al. (2019) for the Western Palaearctic region, and Holloway (1994) for Borneo. More detailed descriptions of taxonomic changes in Ennominae will be given by G. Brehm et al. (2019, unpublished data) and L. MurilloRamos et al. (2019, unpublished data). We here discuss general patterns and give details for taxonomic acts not covered in the other two papers.
Our findings recover Ennominae as a monophyletic entity, but results were not highly supported in RAxML (RBS = 67) compared to IQ-TREE (SH-Like =100, UFBoot2 = 99). The lineage comprising Geometrinae and Oenochrominae is recovered as the sister clade of Ennominae . In previous studies, Wahlberg et al. (2010) sampled 49 species of Ennominae, Õunap et al. (2011) sampled 33 species, and Sihvonen et al. (2011) 70 species including up to eight markers per species. All these studies supported the division of Ennominae into “boarmiine” and “ennomine” moths ( Holloway, 1994). This grouping was proposed by Forbes (1948) and Holloway (1994), who suggested close relationships between the tribes Boarmiini , Macariini, Cassymini and Eutoeini based on the bifid pupal cremaster and the possession of a fovea in the male forewing. The remaining tribes were defined as “ennomines” based on the loss of a setal comb on male sternum A3 and the presence of a strong furca in male genitalia. Both Wahlberg et al. (2010) and Sihvonen et al. (2011) found these two informal groupings to be reciprocally monophyletic.
In our analyses, 653 species with up to 11 markers were sampled, with an emphasis on Neotropical taxa, which so far had been poorly represented in the molecular phylogenetic analyses. Our results recovered the division into two major subclades ( Fig. 6 View Figure 6 ), a core set of ennomines in a well-supported clade, and a poorly supported larger clade that includes the “boarmiines” among four other lineages usually thought of as "ennomines". The traditional “ennomines” are thus not found to be monophyletic in our analyses, questioning the utility of such an informal name. Our phylogenetic hypothesis supports the validation of numerous tribes proposed previously, in addition to several unnamed clades. We validate 23 tribes ( Forum Herbulot, 2007; Skou & Sihvonen, 2015 ): Gonodontini , Gnophini , Odontoperini , Nacophorini , Ennomini , Campaeini , Alsophilini , Wilemaniini, Prosopolophini , Diptychini , Theriini , Plutodini, Palyadini , Hypochrosini , Apeirini , Epionini , Caberini , Macariini, Cassymini , Abraxini , Eutoeini and Boarmiini .
We hereby propose one new tribe: Drepanogynini trib. nov. ( Table 2). Except for the new tribe, most of the groups recovered in this study are in concordance with previous morphological classifications ( Holloway, 1994; Beljaev, 2006, 2016; Forum Herbulot, 2007; Skou & Sihvonen, 2015 ; Müller et al., 2019).
Five known tribes and two further unnamed lineages (E1, E2 in Fig. 6 View Figure 6 ) form the core Ennominae : Gonodontini , Gnophini , Odontoperini , Nacophorini and Ennomini .
Several Neotropical clades that conflict with the current tribal classification of Ennominae will be described as new tribes by G. Brehm et al. (2019, unpublished data). Gonodontini and Gnophini are recovered as sister taxa. Gonodontini was defined by Forbes (1948) and studied by Holloway (1994), who showed synapomorphies shared by Gonodontis Hübner, (1823) , Xylinophylla Warren, 1898 and Xenimpia Warren, 1895 . Our results recovered the genus Xylinophylla as sister of Xenimpia and Psilocladia Warren, 1898 . Psilocladia is an African genus currently unassigned to tribe (see Sihvonen , Staude & Mutanen, 2015 for details). Considering the strong support and that the facies and morphology are somewhat similar to other analyzed taxa in Gonodontini , we formally include Psilocladia in Gonodontini ( Table 2). Gnophini are monophyletic and we formally transfer the African genera Oedicentra Warren, 1902 and Hypotephrina Janse, 1932 , from unassigned to Gnophini ( Table 2). The total number of species, and number of included genera in Gnophini are still uncertain ( Skou & Sihvonen, 2015 ; Müller et al., 2019). Based on morphological examination, Beljaev (2016) treated Angeronini as a synonym of Gnophini . The costal projection on male valva bearing a spine or group of spines was considered as a synapomorphy of the group. Using molecular data, Yamamoto & Sota (2007) showed a close phylogenetic relationship between Angerona Duponchel, 1829 (Angeronini) and Chariaspilates Wehrli, 1953 (Gnophini) . Similar results were shown by Sihvonen et al. (2011) who recovered Angerona and Charissa Curtis, 1826 as sister taxa, and our results also strongly support treating Angeronini as synonym of Gnophini .
Holloway (1994) suggested close affinities among Nacophorini , Azelinini and Odontoperini on the basis of larval characters. In a morphology-based phylogenetic study, Skou & Sihvonen (2015) suggested multiple setae on the proleg on A6 of the larvae as a synapomorphy of the group. Our results also support a close relationship of Nacophorini , Azelinini and Odontoperini . These clades will be treated in more detail by G. Brehmetal . (2019, unpublisheddata).
Following the ideas of Pitkin (2002), Beljaev (2008) synonymized the tribes Ourapterygini and Nephodiini with Ennomini . He considered the divided vinculum in male genitalia and the attachment of muscles m 3 as apomorphies of the Ennomini , but did not provide a phylogenetic analysis. Sihvonen et al. (2011) supported Beljaev’ s assumptions and recovered Ennomos Treitschke, 1825 (Ennomini) , Ourapteryx Leach, 1814 (Ourapterygini) and Nephodia Hübner, 1823 (Nephodiini) as belonging to the same clade. Our comprehensive analysis confirms those previous findings and we agree with Ennomini as the valid tribal name for this large clade. This clade will be treated in more detail by G. Brehm et al.
(2019, unpublisheddata).
Campaeini , Alsophilini , Wilemaniini and Prosopolophini grouped together in a well-supported clade (SH-like = 100, UFBoot2 = 99). Previous molecular analyses have shown an association of Colotoini [= Prosopolophini ] and Wilemaniini ( Yamamoto & Sota, 2007; Sihvonen et al., 2011), although no synapomorphies are known to support synonymization ( Skou & Sihvonen, 2015 ). The Palaearctic genera Compsoptera Blanchard, 1845 , Apochima Agassiz, 1847 , Dasycorsa Prout, 1915 , Chondrosoma Anker, 1854 and Dorsispina Nupponen & Sihvonen, 2013 , are potentially part of the same complex ( Skou & Sihvonen, 2015 , Sihvonen pers. obs.), but they were not included in the current study.
Campaeini is a small group including four genera with Oriental, Palaearctic and Nearctic distribution, apparently closely related to Alsophilini and Prosopolophini , but currently accepted as a tribe ( Forum Herbulot, 2007; Skou & Sihvonen, 2015 ). Our results support the close phylogenetic affinities among these tribes, but due to the limited number of sampled taxa, we do not propose any formal changes.
The genus Declana Walker, 1858 is recovered as an isolated clade sister to Diptychini .
This genus is endemic to New Zealand, but to date has not been assigned to tribe. According to our results, Declana could well be defined as its own tribe. However, the delimitation of this tribe is beyond the scope of our paper and more genera from Australia and New Zealand should first be examined. Aclose relationship between Nacophorini and Lithinini was suggested by Pitkin (2002), based on the similar pair of processes of the anellus in the male genitalia. Pitkin also noted a morphological similarity in the male genitalia (processes of the juxta) shared by Nacophorini and Diptychini . In a study of the Australasian fauna, Young (2008) suggested the synonymization of Nacophorini and Lithinini . This was further corroborated by Sihvonen , Staude & Mutanen (2015) who found that Diptychini were nested within some Nacophorini and Lithinini . However, none of the studies proposed formal taxonomic changes because of limited taxon sampling. In contrast, samples in our analyses cover all biogeographic regions and the results suggest that true Nacophorini is a clade which comprises almost exclusively New World species. This clade is clearly separate from Old World “nacophorines” (cf. Young, 2003) that are intermixed with Lithinini and Diptychini . We here formally transfer Old World nacophorines to Diptychini and synonymize Lithinini syn. nov. with Diptychini ( Table 2). Further formal taxonomic changes in the Nacophorini complex are provided by G. Brehm et al. (2019, unpublished data).
Theria Hübner 1825 , the only representative of Theriini in this study, clustered together with Lomographa Hübner, 1825 ( Baptini in Skou & Sihvonen, 2015 ), in a well-supported clade, agreeing with the molecular results of Sihvonen et al. (2011). The placement of Lomographa in Caberini ( Rindge, 1979; Pitkin, 2002) is not supported by our study nor by that of Sihvonen et al. (2011). The monophyly of Lomographa has not been tested before, but we show that one Neotropical and one Palaearctic Lomographa species indeed group together. Our results show that Caberini are not closely related to the Theriini + Baptini clade, unlike in earlier morphology-based hypotheses ( Rindge, 1979; Pitkin, 2002). Morphologically, Theriini and Baptini are dissimilar, therefore we recognize them as valid tribes (see description and illustrations in Skou & Sihvonen, 2015 ).
According to our results, 11 molecular markers were not enough to infer phylogenetic affinities of Plutodini (represented by one species of Plutodes ). Similar results were found by Sihvonen et al. (2011), who in some analyses recovered Plutodes as sister of Eumelea . Our analyses are congruent with those findings. IQ-TREE results suggest that Plutodes is sister to Palyadini , but RAxML analyses recovered Eumelea as the most probable sister of Plutodes . Given that our analyses are not in agreement on the sister-group affinities of Plutodes , we do not make any assumptions about its phylogenetic position. Instead, we emphasize that further work needs to be done to clarify the phylogenetic positions of Plutodes and related groups.
Hypochrosini is only recovered in a well-defined lineage if the genera Apeira Gistl, 1848 (Apeirini) , Epione Duponchel, 1829 (Epionini) , Sericosema (Caberini) , Ithysia (Theriini) , Capasa Walker, 1866 (unassigned) and Omizodes Warren, 1894 (unassigned) were transferred to Hypochrosini . Skou & Sihvonen (2015) already suggested aclose association of Epionini , Apeirini and Hypochrosini . We think that synonymizing these tribes is desirable. However, due to the limited number of sampled taxa we do not propose any formal changes until more data becomes available. We do suggest, however, formal taxonomic changes for the genera Capasa and Omizodes from unassigned to Hypochrosini ( Table 2).
The southern African genus Drepanogynis is paraphyletic and has earlier been classified as belonging in Ennomini , and later in Nacophorini ( Krüger, 2002) . In our phylogeny, it is intermixed with the genera Sphingomima Warren, 1899 , and Thenopa Walker, 1855 .
Hebdomophruda errans Prout, 1917 also clusters together with these taxa, apart from other Hebdomophruda Warren, 1897 species , which suggests that this genus is polyphyletic.
These genera form a clade sister to the lineage that comprises several Hypochrosini species. Considering that our analysis strongly supports this clade, we place Thenopa , Sphingomima and Drepanogynis in a tribe of their own.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Geometrinae |
|
Tribe |
Archaeobalbini |