
Garra mondica, Sayyadzadeh, Golnaz, Esmaeili, Hamid Reza & Freyhof, Jörg, 2015
|
publication ID |
https://doi.org/10.11646/zootaxa.4048.1.4 |
|
publication LSID |
lsid:zoobank.org:pub:B90AB89E-05ED-44F0-B78C-EBECF5DEC02D |
|
DOI |
https://doi.org/10.5281/zenodo.5614357 |
|
persistent identifier |
https://treatment.plazi.org/id/038F87C5-AF69-FFAB-FF7B-FCADFBB0FCE1 |
|
treatment provided by |
Plazi |
|
scientific name |
Garra mondica |
| status |
sp. nov. |
Garra mondica , new species
( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 )
Holotype. ZM-CBSU H1032, 66 mm SL; Iran: Fars prov.: Konar Siyah spring at Firuzabd, 28°43'40''N 52°25'20''E; G. Sayyadzadeh, M. Razbanian, A. Khajeh Panah and H. Zareiyan, 10 March 2015.
Paratypes. All from Iran. ZM-CBSU H1033, 8, 39–64 mm SL; same data as holotype.—ZM-CBSU K1080, 7, 41–58 mm SL; Iran: Fars prov.: Konar Siyah spring at Firuzabd, 28°43'40''N 52°25'20 ''E; S. Babaee and R. Khaefi, 15 Dec 2012.—ZM-CBSU B242, 1, 48 mm SL; ZM-CBSU B250, 1, 53 mm SL; ZM-CBSU B255, 1, 49 mm SL; Fars prov.: Tang-e-Mohr spring at Lamerd, 27°31'36''N, 52°51'47''E; 6 June 2008.
Additional material. All from Iran. ZM-CBSU B236, 2, 33–47 mm SL; ZM-CBSU B240, 1, 48 mm SL; ZM- CBSU B244, 1, 38 mm SL; ZM-CBSU B246, 4, 39–50 mm SL; ZM-CBSU B251, 4, 37–48 mm SL; Fars prov.: Tang-e-Mohr spring at Lamerd. 27°31'36''N, 52°51'47''E.—ZM-CBSU H1030, 2, 59–63 mm SL; mm SL;—ZM- CBSU H1041, 12, 32–71 mm SL;— FSJF 3488, 3, 53–61 mm SL; Fars prov.: Konar Siyah spring at Firuzabd, 28°43'22''N 52°25'21''E.
Material for molecular genetic analysis: ZM-CBSU M699; Fars prov.: Tang-e-Mohr spring at Lamerd, 27°31'36''N, 52°51'47''E, GenBank accession number: KM214741 View Materials .—ZM-CBSU M793–M794; Fars prov.: Konar Siyah spring at Firuzabd, 28°43'40''N 52°25'20''E, GenBank accession numbers: KM214762 View Materials , KM214736 View Materials .
Diagnosis. Garra mondica is distinguished from all other species of Garra in the rivers flowing to the Persian Gulf south of the Tigris by having a naked breast and anterior belly (vs. fully covered by scales) and a naked middorsal area in front of the dorsal-fin origin or dorsal midline with only few, usually embedded scales ( Fig. 2 View FIGURE 2 ). The naked mid-dorsal area seems to be an unique character state of this species within its comparison group including the species of the Euphrates and Tigris drainages. A naked breast and anterior belly is also found in some populations of G. r u f a. Garra mondica is further distinguished from G. r u f a by having usually 7½ branched dorsalfin rays (vs. usually 8½). Garra mondica is distinguished from all the congeners included in this study by 2 fixed, diagnostic nucleotide substitutions ( Table 1 View TABLE 1 ), and a K2P nearest-neighbor distance of 3.9% to G. rufa ( Table 2 View TABLE 2 ).
Besides the naked breast, anterior belly and predorsal back, G. mondica is also distinguished from G. persica by having usually 9+8 caudal-fin rays (vs. usually 8+8). Garra mondica is distinguished from G. rossica by having a fully-developed mental disc (vs. reduced) and 18–23 total gill rakers on the first branchial arch (vs. 11–15), from G. variabilis by having a fully developed mental disc (vs. reduced) and two pairs of barbels (vs. one) and from G. typhlops and G. widdowsoni by having well-developed eyes and a brown or grey, usually mottled colour pattern (vs. absence of colour pattern).
Nucleotide position relative to Oryzias latipes complete mitochondrial genome ( AP004421 View Materials ).
Nucleotide position relative to Oryzias latipes complete mitochondrial genome ( AP004421 View Materials ).
Description. See Figures 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 for general appearance; morphometric data are given in Table 3 View TABLE 3 . Body elongated, moderately compressed laterally, more compressed in region of caudal peduncle. Dorsal head profile rising gently, slightly convex, more or less continuous with dorsal body profile to nape or about middle between nape and dorsal-fin origin. Ventral profile more or less straight to anal-fin origin. Head moderately large and depressed, with slightly convex or flat interorbital space; height at nape less than head length; width at nape greater or about equal to depth at nape. Snout roundish; transverse lobe with 11–21 tubercles, demarcated posteriorly by a shallow transverse groove in some individuals, no transverse groove in others. Proboscis covered with small or medium sized tubercles, largest on anterior margin of proboscis. Proboscis not or only slightly elevated from depressed rostral surface. Lateral surface of snout covered by small to medium sized tubercles reaching to anterior eye margin in some individuals, or to posterior nostril in others. Depressed rostral surface always without tubercles, anterior arm of depressed rostral surface not reaching to base of rostral barbel, clearly separating transverse lobe from lateral surface. No groove between transverse lobe and lateral surface in some individuals. No head tubercles in juveniles and in a few adults. Eye placed dorso-laterally in posterior half of head. Barbels in two pairs: rostral barbel anterolaterally located, shorter or about equal to eye diameter; maxillary barbel at corner of mouth, shorter than rostral barbel. Rostral cap well-developed, fimbriate, papillate on ventral surface. Upper lip present, as a thin band of papillae arranged in two ridges. Upper jaw almost or completely covered by rostral cap. Disc elliptical, shorter than wide and narrower than head width through base of maxillary barbel; papillae on anterior fold of same size, regularly arranged; groove between antero-median fold and central callous-pad narrow and deep, papillae on inner half of whole length of latero-posterior flap coarsely arranged; anterior marginal surface of central callous pad without or with sparsely arranged small papillae; posterior margin of latero-posterior flap extending vertical to middle of eye.
Dorsal fin with 3 simple and 7½ (28)–8½ (12) branched rays, last simple ray slightly shorter than head length; distal margin concave; origin closer to snout tip than to caudal-fin base; inserted anterior to vertical of pelvic-fin origin; first branched ray longest, tip of last branched ray reaching vertical to, or slightly in front of anus. Pectoral fin with 1 simple and 12–14 branched rays, reaching to a point 3–4 scales anterior to pelvic-fin origin, length shorter than head length. Pelvic fin with 1 simple and 7–8 branched rays, reaching to anus or anal-fin base, origin closer to anal-fin origin than to pectoral-fin origin, inserted below third or fourth branched dorsal-fin ray. Anal fin short, with 3 simple and 5½ branched rays; first branched ray longest; distal margin straight or slightly concave; origin closer to pelvic-fin origin than to caudal-fin base. Anus 2–3 scales in front of anal-fin origin. Caudal fin forked with 9+8 branched rays; tip of lobes pointed. Total gill rakers on first branchial arch 18–23 [18(3), 19(8), 20(6), 21(2), 23(1)]. Lateral line complete, with 28–32 [28(1), 29(2), 30(6), 31(9), 32(8)] scales on body and 2 scales on caudal-fin base. Transverse scale rows above lateral line 3–4; between lateral line and pelvic-fin origin 3 and between lateral line and anal-fin origin 3. Circumpeduncular scale rows 13–14. Usually, no scales on predorsal midline between dorsal-fin origin and nape ( 20 specimens). Seven specimens with 2–4 scales in front of dorsal-fin origin; nine individuals with a few, deeply embedded scales on predorsal midline. Scales on flank regularly arranged. Chest and belly naked. In some individuals belly covered with embedded scales between posterior tip of pectoral fin when folded back and pelvic-fin base. One long axillary scale at base of pelvic fin, 4–6 (usually 5) scales between posteriormost pelvic-fin base and anus, embedded in skin in some individuals.
Coloration. In ethanol: Head, dorsum and flank dark or pale brown. Single, or groups of dark-brown scales on flank. A very faint irregularly shaped midlateral stripe restricted to posterior flank, absent in some individuals, usually interrupted. Mouth, chest and abdomen yellowish white. A wide black or dark-brown bar at posteriormost caudal peduncle, faded in some individuals, bold, 2–3 scales wide in others. Bar reaching dorsal midline in most individuals, not reaching ventral midline. A small black blotch at anterior most lateral line. Lateral line pores cream whitish. Fins hyaline with irregularly set black spots on rays, or rays partly dusty grey or black. Base of last 4 – 6 branched dorsal-fin rays with a black spot or black in individuals larger than 40 mm SL, hyaline in others. In life: Fins hyaline with black spots. Head grey. Flank scales dark grey with individual or groups of pale grey scales forming a mottled pattern; whitish or pale grey on ventral flank and belly. Iris silvery orange. A pale blue dot at anterior-most lateral line reaching down to upper pectoral-fin base in some individuals. A faint irregularly shaped midlateral stripe in few individuals.
Distribution. Garra mondica was found in two small springs in the Mond River drainage in Iran, the Konar Siyah spring ( Fig. 7), which is situated 20 km south-west of Firouzabad and the Tang-e-Mohr spring, which is situated about 10 km south of Mohr.
Etymology. The species is named for the Mond River. An adjective.
Remarks. Garra mondica and G. rufa both occur in the Mond River drainage. While G. r u f a is widespread, it has not been found in sympatry with G. mondica , which has been recorded only from two small springs. Within the Mond River drainage, both species are distinguished by the naked breast, anterior belly and predorsal midline in G. mondica (vs. covered by scales in G. r u f a) and by usually 7½ branched dorsal-fin rays (vs. 8½).
Discognathus crenulatus was described by Heckel (1849) from the area of Shiraz, "Confluenten des Araxes, als aus den Quellen des Saadi und dem Kara-Agatsch [Confluence of Araxes Rivers as well as from the springs of Saadi and from Kara-Agatsch]". The “Confluence of Araxes Rivers” belongs to the Kor drainage, the “springs of Saadi belong to the Lake Mahalo basin and the “Kara-Agatsch” belongs to the Mond River drainage. Coad (1991) treated D. crenulatus as a synonym of G. rufa View in CoL while Bianco & Banarescu (1982) considered it as a subspecies of G. rufa View in CoL . JF examined syntypes of D. crenulatus at NRM (NMW 53236–37). All fishes have the breast and belly as well as the pre-dorsal midline fully covered by scales. No difference between the syntypes of D. crenulatus and G. rufa View in CoL could be found and we follow Coad (1991) and treat D. crenulatus as a synonym of G. rufa View in CoL .
Berg (1949) described G. rufa gymnothorax View in CoL from the Iranian Karun drainage, a tributary of the lowermost Tigris, distinguishing G. r. gymnothorax View in CoL from G. r. r u f a by its naked breast (vs. scaled). Alexander Naseka (Dolsko) examined syntypes (ZIN 13214–15) of this nominal species for us, and these fishes have indeed a naked breast as described by Berg (1949). They are distinguished from G. mondica by having the predorsal midline fully covered by scales (vs. naked in G. mondica ) and usually 8½ branched dorsal-fin rays (vs. usually 7½). Coad (1991) treated G. r. gymnothorax View in CoL as a synonym of G. r u f a. Our molecular data suggest that fishes from the Karun River drainage might represent a distinct species, as they form a separate molecular group, well differentiated from G. r u f a ( Fig. 1 View FIGURE 1 : G. cf. gymnothorax View in CoL ). No Garra View in CoL with a naked breast have been found by us in the Karun River drainage until now. All fishes included in the molecular analysis or examined by us at other occasions have their breast fully covered by scales. While G. gymnothorax View in CoL has been distinguished as a distinct subspecies from G. r u f a by Berg (1949) due to its naked breast, this character seems to be quite variable and does not allow distinguishing our molecular lineage from G. r u f a. The squamation pattern of the breast and belly is quite variable in different populations of G. r u f a and does not allow distinguishing G. mondica from all populations of G. r u f a. Beside the syntypes of G. r. gymnothorax View in CoL , the breast and the midline of the anterior belly are naked in the fishes examined from a tributary to Atatürk reservoir (Euphrates drainage) in Turkey (FSJF 2549). The abdomen is fully covered by embedded, overlapping scales and there are few isolated, deeply embedded scales on the breast in fishes collected from a second tributary to Atatürk reservoir (FSJF 2588). The breast and the anterior abdomen are fully covered by embedded scales except the distalmost pectoral-fin base and the anterior tip of the breast in fishes from the Karun (FSJF 2198), the Karkheh (ZM-CBSU J2501, ZM-CBSU J2404, ZM-CBSU J2084), the Sirvan (ZM-CBSU J2337, ZM-CBSU J2006) and the Zohreh River (ZM-CBSU J1051) and the breast and the anterior abdomen are fully covered by unembedded scales except the anterior tip of the breast, which is naked in fishes from a spring in the Iranian Lake Maharlo basin (FSJF 2238). Additional studies are needed to test whether there are morphological characters distinguishing the molecular lineage called here G. cf. gymnothorax View in CoL from G. r u f a. It is beyond the aims of this study to clarify this issue.
In our molecular dataset, two additional populations of Iranian Garra View in CoL are very closely related to G. mondica . These are fishes from the springs Golabi and Qalatoyeh, both situated in the Kol River drainage in the wider area of the Iranian city of Darab. Fishes from both these populations do not share the diagnostic characters of G. mondica , as both have the breast, anterior belly and predorsal midline covered by scales (vs. naked in G. mondica ) and a greater postorbital distance (40–48% HL vs. in posterior, 36–43% HL). These two populations might also represent G. mondica , which might have a variable squamation pattern as is the case in G. r u f a and the molecular lineage, which might be conspecific with G. gymnothorax View in CoL . But fishes from the Golabi and Qalatoyeh springs are also distinguished from G. mondica by having frequently 8+8 branched caudal-fin rays (vs. almost always 9+8) and frequently 8½ branched dorsal-fin rays (vs. almost always 7½). Therefore, we do not identify these fishes as G. mondica .
As we cannot identify the fishes from the Golabi and Qalatoyeh springs as G. mondica , they might be identified as populations of G. persica View in CoL or G. r u f a that have been introgressed by G. mondica due to postulated past hybridization events. Introgressed fishes can usually be identified as one species, but share the mtDNA with another species. Introgressive hybridization has been documented several times in cyprinid fishes ( Freyhof et al.
2005), but was found to be an uncommon phenomenon in Mediterranean freshwater fishes by Geiger et al. (2014). The springs Golabi and Qalatoyeh are situated in the upper Kol River drainage. Garra persica View in CoL is widespread in the lower Kol River drainage. The fishes from the springs Golabi and Qalatoyeh are distinguished from G. p er s i c a by having 7½ (18) or 8½ (12) branched dorsal-fin rays (vs. 6½ (7)–7½ (10)) and by having frequently 8+8 (5) or 9+8 (22) branched caudal-fin rays (vs. almost always 8+8). If a hybridization of G. mondica with G. persica View in CoL is suspected, this could lead to an increased frequency of fishes with 9+8 branched caudal-fin rays in the hybrid population, as G. mondica has almost always 9+8 and G. persica View in CoL has almost always 8+8 branched caudal-fin rays. This scenario does not explain easily the increased frequency of fishes with 8½ branched dorsal-fin rays. Garra rufa View in CoL is a species widely distributed in all rivers north of the Kol drainage diagnosed by having almost always 8½ branched dorsal-fin rays. While it might be speculated that the fishes from the Golabi and Qalatoyeh springs might have originated from successive hybridization events between G. mondica , G. persica View in CoL and G. r u f a, or might represent a separate, still undescribed species, it is far too speculative and beyond the possibilities of this study to resolve this case. To clarify the evolutionary history of these two populations, in-depth molecular studies are required.
Comparative material. Garra r. gymnothorax : ZIN 13214, 7 syntypes, 39–57 mm SL; Iran: Kulikhan in Karun River drainage, H. Zarudny, 6 June 1904.
Garra cf. gymnothorax : FSJF 2198, 19, 40–94 mm SL; Iran: Kohkeloyeh va Boyrahmad prov.: Beshar River 20 km northeast of Yasooj, 30°44'9.12"N 51°29'31.32"E.— FSJF 2209, 9, 66–129 mm SL; Iran: Chaharmahal Bakhtiari prov.: Stream Sangan at Sangan, 31°15'41.52"N 51°17'9.00"E.
Garra persica : FSJF 2218, 20, 50–83 mm SL; Iran: Hormuzgan prov.: River Rodan about 6 km northeast of Rodan, 27°28'53.70"N 57°15'53.70"E.— FSJF 397, 20, 35–50 mm SL; Iran: Sistan & Baluchestan prov.: Karvander River at bridge north of Karvander, 27°51'18''N 60°46'03''E.—ZM-CBSU 10200, 30, 34–56 mm SL; Iran: Fars prov.: Rasool River at God e Gaz village, 27°17'28.8"N 54°29'20.7"E.
Garra rufa: Qweik drainage: NMW 53240, lectotype ( 101 mm SL) and 7 paralectotypes of D. rufus , 59–101 mm SL; Syria: Aleppo; T. Kotschy, 1842.— NMW 53238, syntypes of D. obtusus , 2, 43– 134 mm SL; Syria: Aleppo; T. Kotschy, 1842. Euphrates and Tigris drainage: NMW 53257, 6 syntypes of D. obtusus , 31–104 mm SL; Iraq: Mossul; T. Kotschy, 1842.— NMW 53236–37, syntypes of D. crenulatus , 4, 56– 74 mm SL; Iran: Qarah Aqaj River.— FSJF 2238, 22, 55–101 mm SL; Iran: Fars prov.: Spring Pirbanoo about 10 km south of Shiraz, 29°31'8.10"N 52°27'55.98"E.— FSJF 2549, 8, 43–82 mm SL; Turkey: Adıyaman prov.: stream Egri south of Adıyaman, a tributary to Atatürk reservoir, 37°44'30.00"N 38°20'6.48"E.— FSJF 2588, 5, 58–65 mm SL; Turkey: Adıyaman prov.: steam Çakal, 13 km west of Adıyaman, tributary to Atatürk reservoir, 37°43'20.53"N 38° 9'55.27"E.— FSJF 2621, 2, 60–67 mm SL; Turkey: Diyabakır prov.: Tigris south of Diyabakır, 37°53'13.80"N 40°13'47.28"E.— FSJF 2849, 7, 71–105 mm SL; Turkey: Diyarbakır prov.: stream Ambar at road to Silvan, 25 km east of Diyarbakır, 37°59'24.72"N 40°22'56.64"E.— FSJF 2864, 3, 98–129 mm SL; Turkey: Batman prov.: Tigris 5 km west of Hasankeyf, 37°43'25.68"N 41°21'37.80"E.— FSJF 2909, 1, 91 mm SL; Turkey: Sivas prov.: stream Kangal under railway bridge at Çetinkaya, 39°15'5.76"N 37°37'8.04"E.— FSJF 2955, 1, 62 mm SL; Turkey: Diyarbakır prov.: stream Savur between Bayındır and Ahmetli east of Tepe, 37°45'49.32"N 40°53'2.04"E.— FSJF 2999, 5, 60–94 mm SL; Turkey: Gaziantep prov.: stream Merziman south of Yavuzeli, 37°17'32.64"N 37°43'23.16"E.— FSJF 3371, 2, 92–100 mm SL; Iraq: stream Suraw near Suraw village, 35°45'45.60"N 45°59'5.40"E.— FSJF 3368, 18, 52–103 mm SL; Iraq: Nalparez River, 35°34'14.40"N 45°51'46.80"E.—ZM-CBSU J 2083, 25, 61–135 mm SL; Iran: Illam prov.: Seimareh River, 33°39'40.3 "N 47°03'29.2 "E.—ZM-CBSU J 2006, 10, 44–78 mm SL; Iran: Kermanshah prov.: Sirvan River, 34°49'37.9 "N 46°21'30 "E.—ZM-CBSU J 1051, 12, 60– 92 mm SL; Iran: Khuzestan prov.: Zohreh River, 30°13'58.56 "N 50°26'50.52 "E. —ZM-CBSU J 2237, 6, 50–89 mm SL; Iran: Kermanshah prov.: Sirvan River, 35°06'43.01 "N 46°15'23.7"E.—ZM-CBSU J 2501, 50, 32–92 mm SL; Iran: Illam prov.: Seimareh River, 33°41'38.1"N 46°42'57.4"E.—ZM-CBSU J2404, 38, 29-92 mm SL; Iran: Illam prov.: Karkheh River, 33°54'26.7"N 46°05'22.8"E.
Garra sp.: FSJF 2255, 22, 52–75 mm SL; ZM-CBSU E812, 87 mm SL; ZM-CBSU E809, 3, 92–125 mm SL; ZM-CBSU E813, 6, 57–84 mm SL; Iran: Fars prov.: Golabi spring about 35 km west of Darab, 28°47'18''N 54°22'15''E.—ZM-CBSU H1056, 20, 62–92 mm SL; Iran: Fars prov.: Rudbal River at Estahban, 28°58'23''N 54°23'25''E; 31 July 2011.—ZM-CBSU H1076, 12, 37–57 mm SL; ZM-CBSU B110, 21, 37–71 mm SL; Iran: Hormuzgan prov.: Haji Abad River about 12 km south-west of Haji Abad, 28°13’33''N 55°49'25''E.—ZM-CBSU H1088, 5, 37–59 mm SL; Iran: Fars prov.: Fadami stream at north of Fadami, 28°13'40''N 55°08'18''E.
Material used in the molecular genetic analysis. Garra barreimiae : FSJF DNA-2488; UAE: Wadi Shawkah, in Emirate of Ras Al-Khaimah, 25°05'54"N 56°06'33"E (GenBank accession numbers: KM214685 View Materials , KM214738 View Materials , KM214758 View Materials , KM214779 View Materials ).— FSJF DNA-2489; UAE: Wadi Wurayah in Emirate of Al-Fujaira, 25°23'54"N 56°16'10"E (GenBank accession numbers: KM214734 View Materials , KM214767 View Materials , KM214783 View Materials ).
Garra cf. gymnothorax : ZM-CBSU M845, M846, M848; Iran: Bashar River, 31°02'25.7"N 51°13'03.6"E (GenBank accession numbers: KM214803 View Materials , KM214735 View Materials , KM214763 View Materials ).—ZM-CBSU M837; Iran: Bashar River, 30°40'50.94"N 51°32'20.72"E (GenBank accession number: KM373234 View Materials ).
Garra cf. longipinnis : FSJF DNA-2493; Oman: Al-Saiq Platue at Jabal al Akhdar, 23°02'00"N 57°28'00"E (GenBank accession numbers: KM214752 View Materials , KM214756 View Materials ).
Garra ghorensis : FSJF DNA-1193; Jordan: Spring at Ghor al Hadithah, 31°17'47"N 35°32'33"E (GenBank accession numbers: KJ553478 View Materials , KJ553508 View Materials , KJ553520 View Materials ).— FSJF DNA-1225; Jordan: stream below Afra hot spring, 30°57'56"N 35°40'56"E (GenBank accession number: KJ553401 View Materials ).— FSJF DNA-2495; Jordan: Wadi Burbaita at the upper part of Wadi Al-Hassa, 30°59'01"N 35°40'11"E (GenBank accession numbers: KM214688 View Materials , KM214728 View Materials , KM214788 View Materials ).— FSJF DNA-2496; Jordan: Wadi Fifa in Southern Ghor, 30°55'52"N 35°28'52"E (GenBank accession number: KM214722 View Materials ).— FSJF DNA-2497; Jordan: Wadi Ibn-Hammad in Al-Karak Provience, 31°18'05"N 35°37'47"E (GenBank accession number: KM214798 View Materials ).
Garra jordanica : FSJF DNA-1186; Jordan: Spring in Suweyma, 31°46'14"N 35°36'10"E (GenBank accession numbers: KM214710 View Materials , KM214750 View Materials ).— FSJF DNA-1206; Syria: River Yarmuk at in Wadi Jallayn, 32°44'21"N 35°58'56"E (GenBank accession number: KJ553525 View Materials ).— FSJF DNA-2498; Jordan: lower most Mujib River, 31°27'22"N 35°35'08"E (GenBank accession numbers: KM214697 View Materials , KM214703 View Materials , KM214753 View Materials , KM214806 View Materials ).
Garra persica : ZM-CBSU M1135, M995; Iran: Rudan River, 27°45'49.4"N 57°14'32.8"E (GenBank accession number: KT808682 View Materials , KM373235 View Materials ).–ZMCBSU M700; Iran: Shur River, 27°19'40.58"N 56°33'10.79"E (GenBank accession number: KM214706 View Materials ). –ZM-CBSU M701; Iran: Kol River, 27°17'28.8"N 54°29'20.7"E (GenBank accession number: KM214807 View Materials ).
Garra rossica : ZM-CBSU M318; Iran: Sistan basin, 34°44'10.1"N 60°34'29.7"E (GenBank accession number: KM214771 View Materials ).—ZM-CBSU M792, M298; Iran: Karvandar River, 27°51'17.97"N 60°46'2.99"E (GenBank accession numbers: KM214796 View Materials , KM214770 View Materials ).—ZM-CBSU M367; Iran: Kavir basin, 32°52'46.74"N 59°3'17.39"E (GenBank accession number: KM214759 View Materials ).
Garra rufa : FSJF DNA-2414; Turkey: Balıklı kaplıca 10 km northeast of Kangal, 39°18'49.07"N 37°28'10.06"E (GenBank accession numbers: KM214792 View Materials - KM214794 View Materials ).— FSJF DNA-923; Turkey: stream Çakal, 13 km west of Adıyaman, tributary to Atatürk reservoir, 37°43'20.36"N 38°9'55.34"E (GenBank accession numbers: KM214692 View Materials , KM214695 View Materials , KM214800 View Materials ).— FSJF DNA-2234; Iraq: Tabin river south of Zarbi, 35°48'19.20"N 45°1'12.00"E.— FSJF DNA-2480; Iraq: Shatt al-Arab at Basrah, 30°32'22"N 47°49"E (GenBank accession number: KM214777 View Materials ).— FSJF DNA-918; Turkey: River Tigris south of Diyabakır, 37°53'13.80"N 40°13'47.40"E (GenBank accession number: KM214684 View Materials ). –ZM-CBSU M849, M850; Iran: Dalaki River, 29°28'15.40"N 51°18'14.60"E (GenBank accession: KT808680 View Materials , KT808681 View Materials ).—ZM-CBSU M704, M836; Iran: Koohmareh Sorkhi River at Helleh, 29°23'39.80"N 52°9'40.20"E (GenBank accession numbers: KM214760 View Materials , KM214810 View Materials ).—ZM-CBSU M847; Iran: Bashar River at Yasouj, 31° 2'25.68"N 51°13'3.60"E (GenBank accession number: KM214711 View Materials ).—ZM-CBSU M745; Iran: Doirej River at Ilam, 32°39'13.13"N 47°32'36.97"E (GenBank accession number: KM214733 View Materials ).—ZM-CBSU M713; Iran: Kavar River at Mond, 29°10'54.60"N 52°41'28.20"E (GenBank accession number: KM373232 View Materials ). –ZM-CBSU M842; Iran: stream at Saadi Tomb, 29°37'20.77"N 52°34'55.59"E (GenBank accession number: KM214714 View Materials ).—ZM-CBSU M711; Iran: Fahlian River, 30°11'4.40"N 51°31'25.57"E (GenBank accession number: KM214709 View Materials ).
Garra sp.: ZM-CBSU M299, M301, M302; Fars prov.: Golabi spring about 35 km west of Darab, 28°47'18''N 54°22'15''E (GenBank accession numbers: KM214691 View Materials , KM214782 View Materials , KM214791 View Materials ).
Garra typhlops : FSJF DNA-2169; Iran: Well at Kaaje-Ru, 33°04'39"N 48°35'33"E (GenBank accession numbers: KM214717 View Materials , KM214731 View Materials ).
Garra variabilis : FSJF DNA-1159; Syria: Nahr al Barid at Nahr al Barid, 35°18'08"N 36°20'43"E (GenBank accession numbers: KJ553391 View Materials , KJ553567 View Materials )— FSJF DNA-1168; Syria: Orontes at Shayzar, 35°16'18"N 36°33'46"E (GenBank accession numbers: KJ553422 View Materials , KJ553524 View Materials ).
Garra widdowsoni : FSJF DNA-2301; Iraq: cavern 25 feet below ground, 6 miles north of Haditha, 34° 4'0.00"N 42°23'60.00"E (GenBank accession numbers: KM214769 View Materials , KM214795 View Materials ).
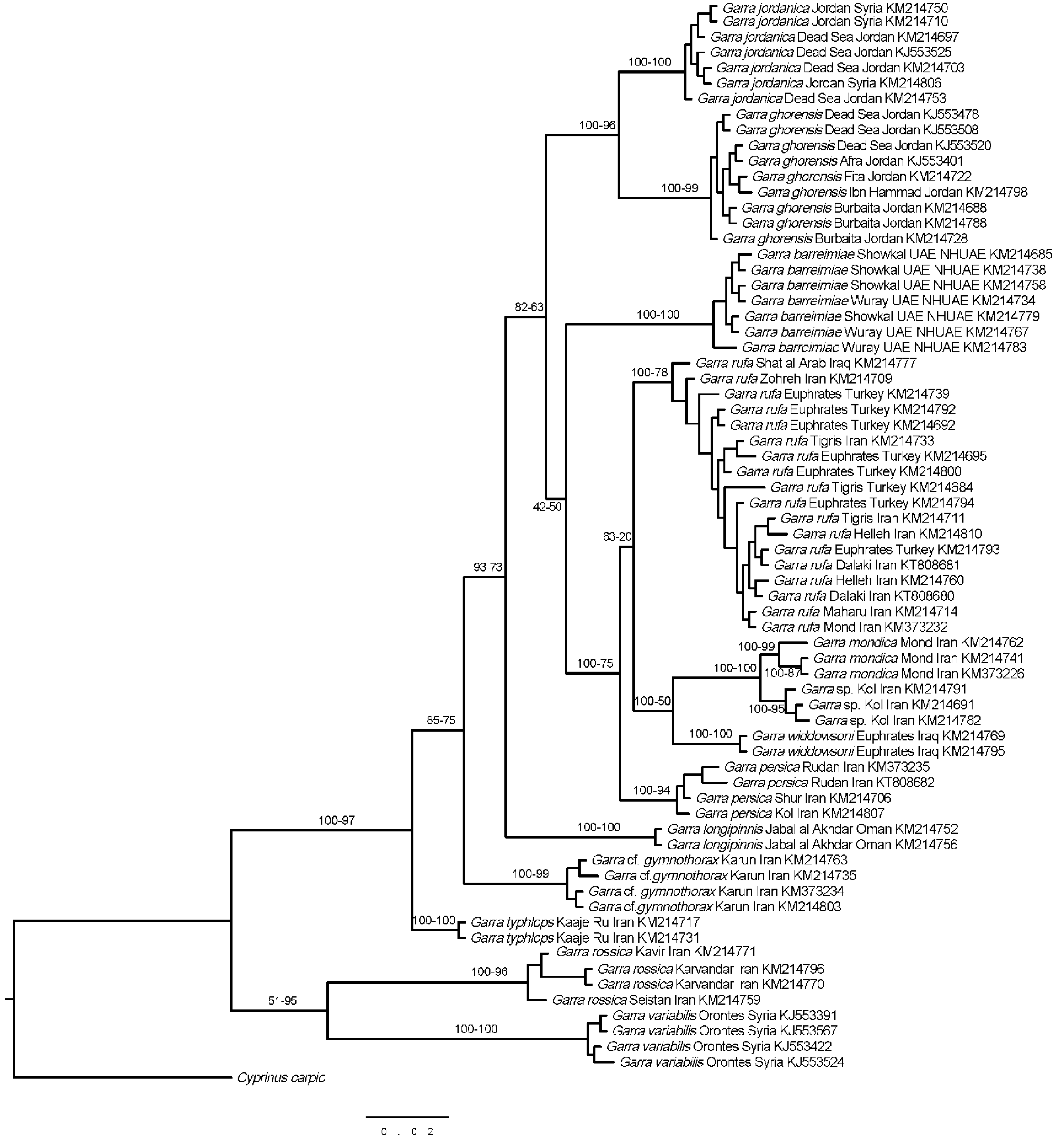



TABLE 1. Diagnostic nucleotide substitutions found in the COI barcode region of Garra species studied.
| Nucleotide position | N | 5 9 2 8 | 5 9 3 4 | 5 9 4 3 | 5 9 4 6 | 5 9 7 3 | 5 9 7 9 | 5 9 9 1 | 6 0 0 0 | 6 0 0 3 | 6 0 0 6 | 6 0 0 9 | 6 0 2 7 | 6 0 3 3 | 6 0 3 4 | 6 0 3 6 | 6 0 4 0 | 6 0 5 1 | 6 0 5 4 | 6 0 5 8 | 6 0 6 0 | 6 0 7 5 | 6 0 7 8 | 6 1 1 7 | 6 1 2 6 | 6 1 3 2 | 6 1 4 1 | 6 1 4 4 | 6 1 4 7 | 6 1 5 4 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| G. barreimiae | 7 | T | A | T | T | C | C | T | A | C | C | G | C | A | C | A | C | G | A | C | A | A | G | C | A | A | T | T | T | C |
| G. ghorensis | 9 | T | G | T | T | C | C | T | A | A | C | G | C | A | C | A | C | G | G | C | A | A | G | C | A | A | T | T | T | C |
| G. jordanica | 7 | T | G | T | T | C | C | T | A | C | C | G | C | A | C | A | C | G | A | C | A | A | G | C | A | A | T | T | T | C |
| G. cf. longipinnis | 2 | T | G | T | T | C | C | T | A | C | C | G | C | A | T | A | T | G | A | C | G | A | G | C | A | A | T | T | T | C |
| G. mondica | 3 | T | G | C | T | C | C | C | A | C | C | G | C | A | C | A | C | A | A | C | A | A | G | C | A | A | T | T | T | C |
| G. persica | 4 | T | G | T | T | C | C | T | A | C | C | A | C | A | C | A | C | A | A | C | A | A | G | C | A | A | T | T | T | C |
| G. rossica | 4 | T | G | T | T | C | C | T | T | T | C | A | T | G | C | A | C | T | C | T | A | A | A | C | A | T | T | C | C | T |
| G. r u f a | 18 | T | G | T | T | C | C | T | A | C | C | G | C | A | C | A | C | A | A | C | A | A | G | C | A | A | T | T | T | C |
| G. typhlops | 2 | T | G | T | T | C | C | T | A | C | C | G | C | A | C | A | C | G | T | C | A | A | G | C | A | A | T | T | T | C |
| G. variabilis | 4 | T | G | T | C | T | T | T | C | T | T | T | C | T | C | A | C | A | C | C | A | A | G | C | G | C | T | T | T | C |
| G. widdowsoni | 2 | C | G | T | T | C | C | T | A | C | C | G | C | A | C | A | C | A | A | C | A | G | G | C | A | A | C | T | T | C |
| G. cf. gymnothorax | 4 | T | G | T | T | C | C | T | A | C | C | G | C | A | C | G | C | G | A | C | A | A | G | T | A | A | T | T | T | C |
| continued. |
TABLE 2. Estimates of evolutionary divergence (%) over sequence pairs between species found in the COI barcode region of Garra species studied.
| G. r u f a | N 18 | rufa . G | mondica . G | Kol. sp. G | persica . G | gymnothorax . cf. G | typhlops . G | widdowsoni . G | jordanica . G | barreimiae . G | longipinnis ghorensis . cf.. G G | a c i s s ro. G |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| G. mondica | 3 | 3.88 | ||||||||||
| G. sp. Kol | 3 | 3.51 | 0.73 | |||||||||
| G. persica | 4 | 2.43 | 4.13 | 3.8 | ||||||||
| G. cf. gymnothorax | 4 | 5.67 | 5.29 | 5.8 | 5.48 | |||||||
| G. typhlops | 2 | 5.12 | 6.31 | 6.08 | 4.77 | 4.49 | ||||||
| G. widdowsoni | 2 | 3.17 | 4.05 | 3.69 | 3.8 | 7.16 | 6.99 | |||||
| G. jordanica | 7 | 4.15 | 6.16 | 5.71 | 5.26 | 6.32 | 6.15 | 6.39 | ||||
| G. barreimiae | 7 | 6.67 | 7.46 | 7.68 | 6.22 | 7.39 | 7.45 | 7.69 | 7.3 | |||
| G. cf. longipinnis | 2 | 7.09 | 6.97 | 6.74 | 6.19 | 6.68 | 5.4 | 8.12 | 7.73 | 7.66 | ||
| G. ghorensis | 9 | 5.57 | 7.06 | 6.75 | 5.98 | 7.6 | 6.52 | 7.45 | 4.39 | 8.37 | 7.42 | |
| G. rossica | 4 | 13.6 | 13.7 | 13.4 | 13 | 14.1 | 11.5 | 15 | 14.7 | 15.3 | 13.9 13.69 | |
| G. variabilis | 2 | 13.8 | 16.3 | 15.7 | 12.7 | 14.6 | 11.9 | 15.5 | 13.9 | 17 | 13.6 13.52 | 11.43 |
TABLE 3. Morphometric data of Garra mondica holotype ZM-CBSU H 1032, paratypes, ZM-CBSU K 1079 - K 1086, ZM-CBSU H 1033 - H 1040, ZM-CBSU B 242, ZM-CBSU B 250, ZM-CBSU B 255, n = 20.
| Holotype | Range | Mean | SD | |
|---|---|---|---|---|
| Standard length (mm) | 65.9 | 38.5–65.9 | ||
| In percent of standard length | ||||
| Head length | 25.7 | 24.0–27.4 | 25.9 | 1.0 |
| Body depth at dorsal-fin origin | 18.5 | 16.2–23.4 | 18.5 | 2.0 |
| Predorsal length | 47.5 | 46.1–50.3 | 48.2 | 1.1 |
| Postdorsal length | 40.2 | 37.4–41.9 | 39.6 | 1.5 |
| Preanal length | 72.0 | 71.1–74.6 | 73.1 | 1.0 |
| Prepelvic length | 51.5 | 49.6–55.3 | 52.8 | 1.5 |
| Distance between pectoral and pelvic-fin origins | 30.0 | 28.2–31.6 | 30.0 | 1.1 |
| Distance between pelvic and anal-fin origins | 19.5 | 17.4–21.6 | 19.6 | 1.1 |
| Depth of caudal peduncle | 11.1 | 10.5–12.4 | 11.6 | 0.6 |
| Length of caudal peduncle | 20.6 | 18.8–22.2 | 20.4 | 0.8 |
| Dorsal-fin base length | 13.6 | 12.0–15.1 | 13.2 | 0.9 |
| Anal-fin base length | 8.4 | 6.9–8.4 | 7.5 | 0.4 |
| Pectoral-fin length | 24.3 | 23.0–27.7 | 24.8 | 1.4 |
| Pelvic-fin length | 20.1 | 18.1–21.3 | 19.8 | 0.9 |
| In percent of head length | ||||
| Head depth at eye | 59 | 55–66 | 59.8 | 4.1 |
| Snout length | 47 | 38–47 | 43.0 | 2.6 |
| Postorbital distance | 40 | 36–43 | 39.5 | 1.9 |
| Interorbital width | 40 | 37–46 | 41.8 | 2.3 |
| Eye diameter | 15 | 13–22 | 17.8 | 2.5 |
| Maximum head width | 68 | 66–74 | 68.6 | 2.3 |
| Rostral barbel length | 18 | 16–20 | 17.9 | 1.5 |
| Maxillary barbel length | 13 | 12–17 | 14.5 | 1.1 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |