
Ichnanthus adpressus C. Silva & R.P. Oliveira, 2013
|
publication ID |
https://doi.org/ 10.11646/phytotaxa.104.1.3 |
|
persistent identifier |
https://treatment.plazi.org/id/03905075-647B-AB74-FF00-FBEEBF54FB18 |
|
treatment provided by |
Felipe |
|
scientific name |
Ichnanthus adpressus C. Silva & R.P. Oliveira |
| status |
sp. nov. |
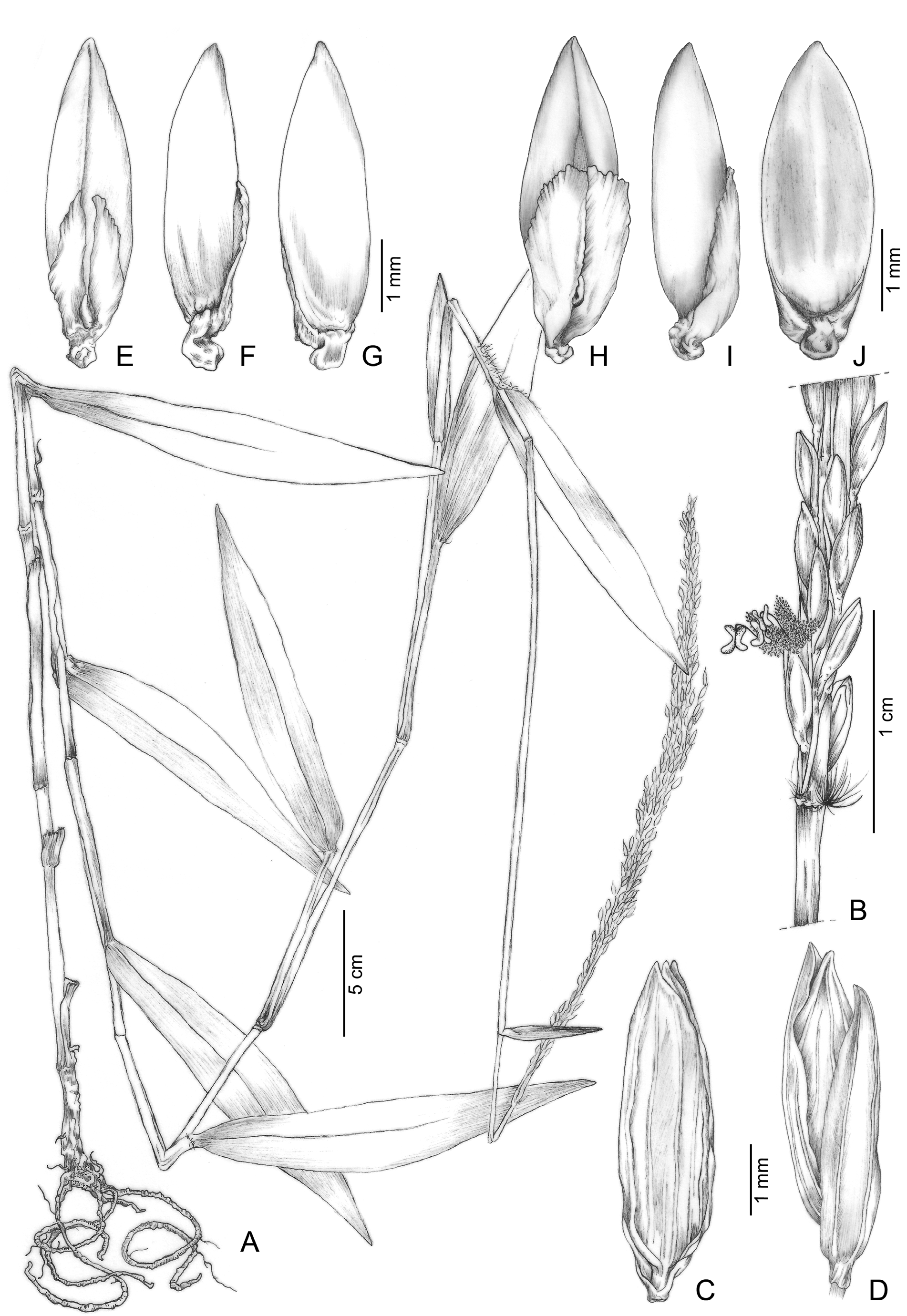
Ichnanthus adpressus C. Silva & R.P. Oliveira View in CoL , sp. nov. ( Figs. 1−3 View FIGURE 1 View FIGURE 2 View FIGURE 3 )
It differs from other species with winged appendages by the combination of caespitose habit with decumbent flowering culms, sometimes scandent, leaves distributed along the culm, contracted paniculate inflorescences with branches appressed to the main axis and fleshy, translucent winged appendages with short filiform epidermal projections.
Type: — BRAZIL. Minas Gerais: Lima Duarte, Parque Estadual de Ibitipoca , ca. 1370 m, 21°42’31”S, 43°53’45”W, 23 July 2012 (fl,fr), C. Silva & C. Snak 993 (holotype HUEFS!, isotypes RB!, ICN!, K!, NY!, P!, CESJ!, CANB!) GoogleMaps
Perennial plants, herbaceous, caespitose, without rhizomes, 0.2−1.5 m tall. Culms erect, the flowering ones elongated and decumbent, sometimes scandent, rooting at the lower nodes; internodes glabrous or sparsely pilose; nodes glabrous or pilose. Leaves distributed along the culms; leaf sheaths pilose at the base, margins and apex, or densely hirsute; collar pilose, trichomes 0.5−2.5 mm; leaf blades 3−16 × 0.5−2.9 cm, ovatelanceolate to elliptic-lanceolate, acute to acuminate, rounded at the base, glabrous on both surfaces, bearing trichomes only along the margins on the abaxial surface, on the lower 1/3 of the blade, or sparsely to densely pilose on both surfaces; midrib inconspicuous adaxially and slightly pronounced abaxially, transverse crossveins conspicuous abaxially (tessellation), marginal rib slightly scabrous; pseudopetioles absent; ligules membranous-ciliate, 1−2 mm long (ca. 0.2 mm membranous). Inflorescence paniculate, contracted, with branches appressed to the main axis, 11−55 (5.5−31.5 cm exserted) × 1−2 cm, terminal, partially included to entirely exserted from the terminal leaf sheath; primary branches 1.8−10.4 cm long, sub-verticillate to alternate toward the apex, scabrous, the lower ones longer than the apical ones; secondary branches 1.5−3.8 cm, scabrous, only at the lower primary branches; spikelets in pairs, the lower ones with pedicels 1.8−4 mm long, and the apical ones with pedicels 4−9.5 mm long, scabrous. Spikelets 4−5.2 × 1−1.5 mm, ellipticlanceolate to oblong-lanceolate, regularly distributed along the branches, greenish, vinaceous at margins toward the apex, glabrous; internode between glumes conspicuous, ca. 0.5 mm long; glumes subequal, the lower one 3.8−5 mm long, 3-nerved, margins hyaline, apex acute; the upper one 3.5−4.8 mm long, 5-nerved, margins hyaline, apex acute; lower anthecium sterile; lemma 4−5 mm long, 5-nerved, margins hyaline, apex acute; palea 3/4 length of the lemma, hyaline; upper anthecium 3.8−4.5 × 1−1.2 mm, ovate-lanceolate, pale to brownish with dark spots at maturity, glabrous, smooth; rachilla appendages 1.7−2.2 mm long, winged, fleshy and translucent, 1/2−3/5 the length of anthecium, with short filiform epidermal projections along its surface, apex acute to rounded at maturity, asymmetrical, converging towards the mid portion of the anthecium, pale to slightly orange, emerging from a curved stipe at the base of the upper lemma, 0.3−0.6 mm long; palea with an acute protruding base conspicuous on the young anthecium, inconspicuous at maturity; stamens 3, filaments whitish, anthers purplish; stigmas 2, vinaceous. Caryopsis 2 × 1 mm, oblong, pale brown; hilum punctiform.
Etymology: —From the Latin words ad = towards; presso = press; in reference to the contracted paniculate inflorescences with branches appressed to the main axis ( Clifford & Bostock 2007), uncommon among species of Ichnanthus which usually have open paniculate inflorescences.
Similarities to other species of Ichnanthus : —In combination, the habit ( Fig. 2A View FIGURE 2 ) and inflorescence ( Fig. 2B,C View FIGURE 2 ) of I. adpressus distinguish it from other known species of Ichnanthus . Contracted paniculate inflorescences with appressed branches can also be found in I. riedelii ( Trinius 1834: 327) Döll (1877: 277) , but it differs from the new species by having predominantly basal leaves with blades which are long-attenuate at the base ( Boechat 2005) instead of leaves distributed along the culm with blades which are rounded at the base.
The spikelets of the new species ( Fig. 2E View FIGURE 2 ) are very similar in size, texture and proportions of its parts to those of Ichnanthus lancifolius Mez (1918: 126) , but the latter can be distinguished by its open paniculate inflorescences ( Boechat 2005). Similarly to I. riedelii , I. lancifolius can also be distinguished from I. adpressus by having predominantly basal leaves, although in this species the blades may be short- or longattenuate at the base ( Boechat 2005).
According to Boechat’s (2005) key to the species of Ichnanthus from Brazil, Ichnanthus adpressus could be placed in the group composed of I. nemoralis ( Schrader 1824: 255) Hitchcock & Chase (1917: 334) , I. inconstans (Trinius ex Nees 1829: 132) Döll (1877: 284) , I. hirtus ( Raddi 1823: 365) Chase (1923: 175) , I. dasycoleus Tutin (1934: 337) , I. leiocarpus and I. calvescens (Nees ex Trinius 1826: 193) Döll (1877: 285) , due to the presence of winged appendages, leaves that are distributed along the culms, a muticous lower glume, a glabrous upper lemma, one single terminal inflorescence and blades larger than 7 × 1 cm. All these species display open paniculate inflorescences, an upper palea which is rounded at the base, and surface of the winged appendages without epidermal projections (C. Silva et al., unpublished data), unlike the contracted paniculate inflorescences, an upper palea with an acute protuberance at the base, and surface of the winged appendages with filiform epidermal projections in I. adpressus .
In addition, some specimens of the new species with teratogenic spikelets were observed in the materials collected in the field as well as in herbarium material (e.g. R.C. Oliveira 20, P.L. Viana et al. 2149). These anomalous spikelets are characterized by the proliferation of anthecia, making them multiflowered and falciform ( Fig. 2D View FIGURE 2 ). These have also been mentioned for I. inconstans ( Oliveira et al. 2003, Boechat 2005, Dias-Melo et al. 2009) but not for other species of Ichnanthus .
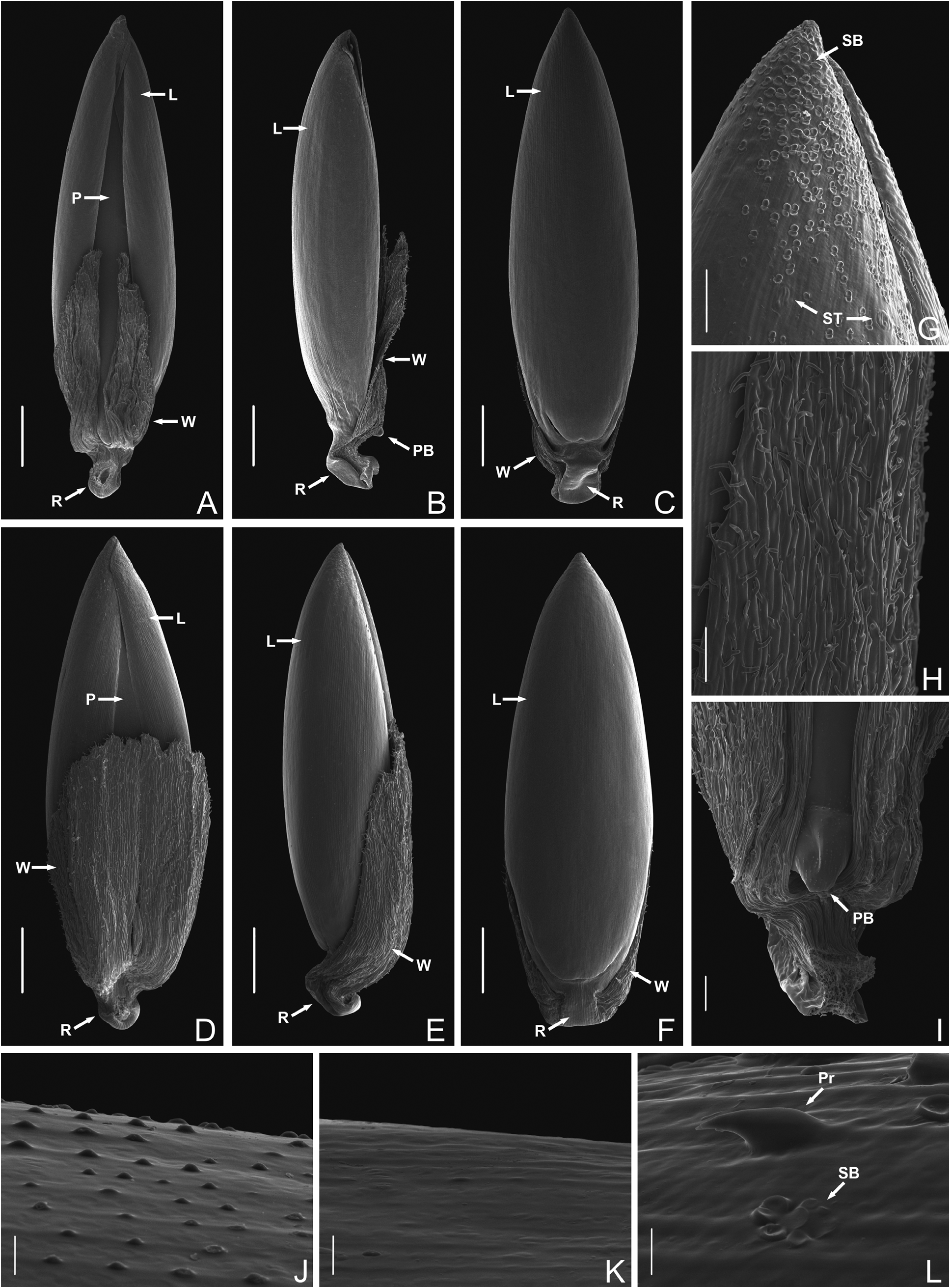
Micromorphology of the upper anthecium: —The upper anthecium of I. adpressus has winged appendages, which change shape throughout its development at the same time as the change in the lemma and palea surfaces ( Fig. 1 View FIGURE 1 E−J and 3A−F). The winged appendages of the young anthecium are narrower and plicate ( Fig. 1 View FIGURE 1 E−G and 3A−C), becoming larger and inflated at maturity ( Fig. 1 View FIGURE 1 H−J, 2F,G and 3D−F). According to Shaw & Webster (1983) the plicate surface of the winged appendages could be “the result of dehydration”, but our data suggest that it is a matter of the stage of maturation, in this case a young stage. The surfaces of the lemma and the palea also change, being slightly papillate (rounded papillae 2−7.5 µm wide) on the young anthecium ( Fig. 3J View FIGURE 3 ) with the papillae being no longer visible on the mature anthecium ( Fig. 3K View FIGURE 3 ). The presence of papillae has only been previously cited for the palea in Ichnanthus ( Shaw & Webster 1983) . The papillae of I. adpressus are smaller than those of other species of Ichnanthus mentioned by Shaw & Webster (1983, 10− 25 µm wide), and smaller than those of I. longhi-wagnerae Mota & Oliveira (2012: 117 ; 5−8 µm wide).
The winged appendages arise from a curved rachilla that subtends the upper anthecium ( Fig. 3B,E View FIGURE 3 ) as indicated by Shaw & Webster (1983) for I. panicoides Beauvois (1812: 56) , I. leptophyllus Döll (1877: 287) and I. longiglumis Mez (1918: 131) . The surface is covered with numerous short unicellular macrohairs, 12−49 µm long ( Fig. 3H View FIGURE 3 ), previously recorded only for I. leptophyllus ( Shaw & Webster 1983) . The palea possesses a protruding base ( Fig. 3B,I View FIGURE 3 ), which has been recorded for species with scars or swellings ( Shaw & Webster 1983) but not yet for species with winged appendages.
The apex of the lemma has numerous silica-bodies on its surface ( Fig. 3G View FIGURE 3 , 12−26 µm wide), which Shaw & Webster (1983) called “exfoliated silica-bodies” in Echinolaena standleyi ( Hitchcock 1930: 662) Stieber (1987: 214) . There are also a few prickle hairs ( Fig. 3L View FIGURE 3 ) and stomata ( Fig. 3G View FIGURE 3 ) on the surface of the lemma. Thus, the presence of macrohairs on the surface of the winged appendages, silica-bodies on the surface of the apex of the lemma, and papillae and prickle hairs on the lemma characterize I. adpressus . Furthermore, the latter two features are recorded for the first time in a lemma of an Ichnanthus species.
Life history and phenology: —The populations of I. adpressus are somewhat abundant in the cloud forests of the PEIB and SN and the individuals grow close to each other in small clumps ( Fig. 2A View FIGURE 2 ); some are more robust and taller than others ( Fig. 1A View FIGURE 1 ). The clump starts with a few culms growing obliquely to the ground, then these culms ascend and the internodes elongate. Generally, the flowering culms become longer than the vegetative ones and grow prostrate on the surrounding vegetation.
We collected the new species with flowers and fruits in March, July and December and we also observed it flowering in January at the PEIB. Amongst the herbarium specimens analyzed there are collections from every month of the year except September; the new species is probably fertile throughout the year.
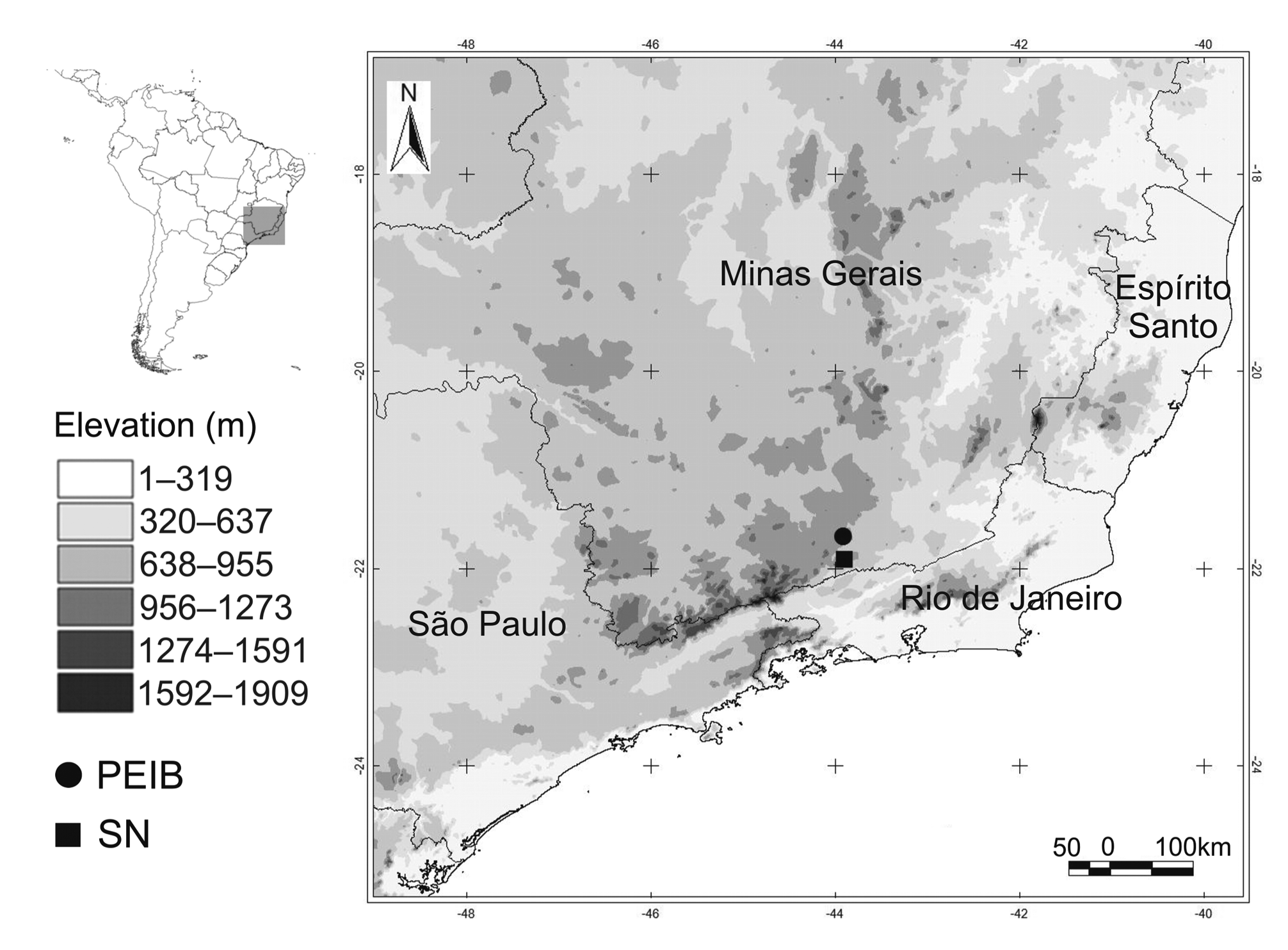
Distribution and habitat: — Ichnanthus adpressus is only known from two areas in southern Minas Gerais, the PEIB and the SN ( Fig. 4 View FIGURE 4 ), both situated at the ranges of the Serra da Mantiqueira within the Atlantic Rain Forest domain. The first is partly located at the municipality of Santa Rita de Ibitipoca and partly at the municipality of Lima Duarte, and the latter is located between the municipalities of Rio Preto, Lima Duarte, Santa Bárbara do Monte Verde and Olaria. These two areas have a peculiar vegetation type, composed of a mosaic of grassland, scrub and cloud forest associated with quartzite rock outcrops, at 1200−1800 m elevation. Quartzite rock outcrops are unusual in the Serra da Mantiqueira where rocky vegetation is mainly associated with granite and gneiss outcrops ( Stafford 1999). Both areas were listed as conservation priority for their high fauna and flora species richness and endemism ( Drummond et al. 2005). Another grass species, Chusquea riosaltensis Clark (1992: 403) , possess a distribution pattern similar to that of I. adpressus , known only from the PEIB and the SN ( Moreira et al. 2008).
The new species is found at the border and inside the cloud forests (also called “Mata baixa com Candeia” by Dias et al. 2002). These forests are composed of small spaced trees with a dense cover of lichen providing a partially shaded understory. The ground is densely covered by mosses and has a high content of organic matter.
Conservation status: —Despite the fact that several collections of I. adpressus are known, its occurrence is restricted to the PEIB and the SN, which fits the IUCN Red List Criteria B2ab(ii) ( IUCN 2012). According to these criteria, I. adpressus is here assessed as Endangered (EN). Furthermore, although the PEIB is under protection by the government of Minas Gerais, the SN is not protected and includes several private properties, threatening the conservation of the area. Moreover, the cloud forests represent only a small portion of these two areas.
Additional specimens examined (paratypes): — BRAZIL. Minas Gerais: Lima Duarte , 6 February 1989 (fl,fr), L . Krieger & Leise s.n. ( CESJ!, HURG, MBM, BHCB, ESA); 6 February 1989 (fl,fr), L . Krieger & L. T . Souza s.n. ( RB!, CESJ); Lima Duarte-Olaria, 27 March 1989 (fl,fr), L . Krieger & M . Brügger s.n. ( CESJ!, HURG); Lima Duarte, Parque Estadual de Ibitipoca ( PEIB), nascente do Rio Pirapitinga , 11 October 1989 (fl,fr), L . Krieger & M . Brügger s.n. ( CESJ!, HURG); ibid, 15 August 1990 (fl,fr), M . Brügger 683 ( CESJ!); PEIB , aceiro inferior, 20 June 1991 (fl,fr), R. C . Oliveira 19 ( CESJ!, HURG); PEIB , estrada para o alojamento, 20 June 1991 (fl,fr), R. C . Oliveira 20 ( CESJ!, CEN, HURG); PEIB , Prainha , 22 February 1992 (fl,fr), M . Eiterer s.n. ( CESJ!); PEIB , Ponte de Pedra , 22 February 1992 (fl,fr), M . Eiterer et al. s.n. ( CESJ!); PEIB , próximo à trilha para a Cachoeira dos Macacos , 19 October 2003 (fl,fr), R . Dias-Melo et al. 15 ( RB!); PEIB , próximo à entrada do Parque , 21 December 2003 (fl,fr), F. M . Ferreira & L. M . Neto 614 ( RB!, CESJ); PEIB , atrás da casa da Polícia Florestal , 4 February 2004 (fl,fr), R . Dias-Melo et al. 138 ( RB!, K); PEIB , estrada para a ponta do Parque , 5 February 2004 (fl,fr), R . Dias-Melo et al. 158 ( RB!); PEIB , trilha Lanchonete-Ponte de Pedra , 31 March 2004 (fl,fr), R. C . Forzza et al. 3296 ( RB!); PEIB , trilha para a Cachoeirinha , Gruta dos Três Arcos, 31 March 2004 (fl,fr), R . Dias-Melo et al. 182 ( RB!, K, MBM, SP); PEIB , mata em frente ao alojamento, 17 March 2005 (fl,fr), R . Marquete & E . von S. Medeiros 3583 ( RB!); PEIB , Ponte de Pedra , 26 May 2005 (fl,fr), R. C . Forzza et al. 3983 ( RB!, SP, K); PEIB , trilha para o Pico do Peão, próximo ao Monjolinho , 9 August 2005 (fl,fr), R . Dias-Melo et al. 294 ( RB!, K, MBM, SP); PEIB , Norte do Parque , 7 March 2006 (fl,fr), F. M . Ferreira et al. 981 ( CESJ!); PEIB , mata ciliar próximo ao alojamento, 17 March 2009 (fl,fr), L. L . Giacomin et al. 728 ( BHCB!); PEIB , mata nos arredores da Prainha , 21°42’29”S, 43°53’36”W, 1355 m elev., 24 July 2012 (fl,fr), C GoogleMaps . Silva & C . Snak 994 ( HUEFS!); PEIB , trilha do Mirante, na altura do Lago das Miragens , 21°42’37”S, 43°53’35”W, 1343 m elev., 24 July 2012 (fl,fr), C GoogleMaps . Silva & C . Snak 997 ( HUEFS!, CEN!, INPA!, MG!); PEIB , mata na trilha entre o Mirante do Gavião e a Ponte de Pedra , 21°42’51”S, 43°53’39”W, 24 July 2012 (fl,fr), C GoogleMaps . Silva & C . Snak 999 ( HUEFS!, MBM!, R!, RBR!, SP!); Rio Preto, Serra Negra ( SN), trilha para o Ninho da Égua , 21 August 2004 (fl,fr), C. N . Matozinhos et al. 53 ( CESJ!); SN , trilha para o Ninho da Égua , 9 November 2005 (fl,fr), C. N . Matozinhos et al. 242 ( CESJ!, HUEFS!); SN , subida para a Cachoeira Ninho da Égua , 1 July 2006 (fl,fr), P. L . Viana et al. 2149 ( CESJ!, HUEFS!); SN , Serrinha, Sítio do Rinaldo Degredo , 28 July 2009 (fl,fr), F. S . Souza et al. 756 ( CESJ!) .

| C |
University of Copenhagen |
| HUEFS |
Universidade Estadual de Feira de Santana |
| RB |
Jardim Botânico do Rio de Janeiro |
| ICN |
Instituto de Ciencias Naturales, Museo de Historia Natural |
| K |
Royal Botanic Gardens |
| NY |
William and Lynda Steere Herbarium of the New York Botanical Garden |
| P |
Museum National d' Histoire Naturelle, Paris (MNHN) - Vascular Plants |
| CESJ |
Universidade Federal de Juiz de Fora |
| CANB |
Australian National Botanic Gardens |
| L |
Nationaal Herbarium Nederland, Leiden University branch |
| HURG |
Universidade do Rio Grande (FURG) |
| MBM |
San Jose State University, Museum of Birds and Mammals |
| BHCB |
Universidade Federal de Minas Gerais |
| ESA |
Universidade de São Paulo |
| T |
Tavera, Department of Geology and Geophysics |
| M |
Botanische Staatssammlung München |
| R |
Departamento de Geologia, Universidad de Chile |
| CEN |
EMBRAPA Recursos Geneticos e Biotecnologia - CENARGEN |
| F |
Field Museum of Natural History, Botany Department |
| SP |
Instituto de Botânica |
| E |
Royal Botanic Garden Edinburgh |
| INPA |
Instituto Nacional de Pesquisas da Amazonia |
| MG |
Museum of Zoology |
| RBR |
Universidade Federal Rural do Rio de Janeiro |
| SN |
South China Normal University |
| N |
Nanjing University |
| S |
Department of Botany, Swedish Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |