
Microhyla aurantiventris, Nguyen & Poyarkov & Nguyen & Nguyen & Tran & Gorin & Murphy & Nguyen, 2019
|
publication ID |
https://doi.org/10.11646/zootaxa.4543.4.4 |
|
publication LSID |
lsid:zoobank.org:pub:6D132076-BE52-4C17-91A5-1D372CD8CA29 |
|
DOI |
https://doi.org/10.5281/zenodo.5937224 |
|
persistent identifier |
https://treatment.plazi.org/id/0390521C-064F-5854-FF26-FE6BFC970C2F |
|
treatment provided by |
Plazi |
|
scientific name |
Microhyla aurantiventris |
| status |
sp. nov. |
Microhyla aurantiventris View in CoL sp. nov.
Figs. 3–7 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 ; Tables 4, 5.
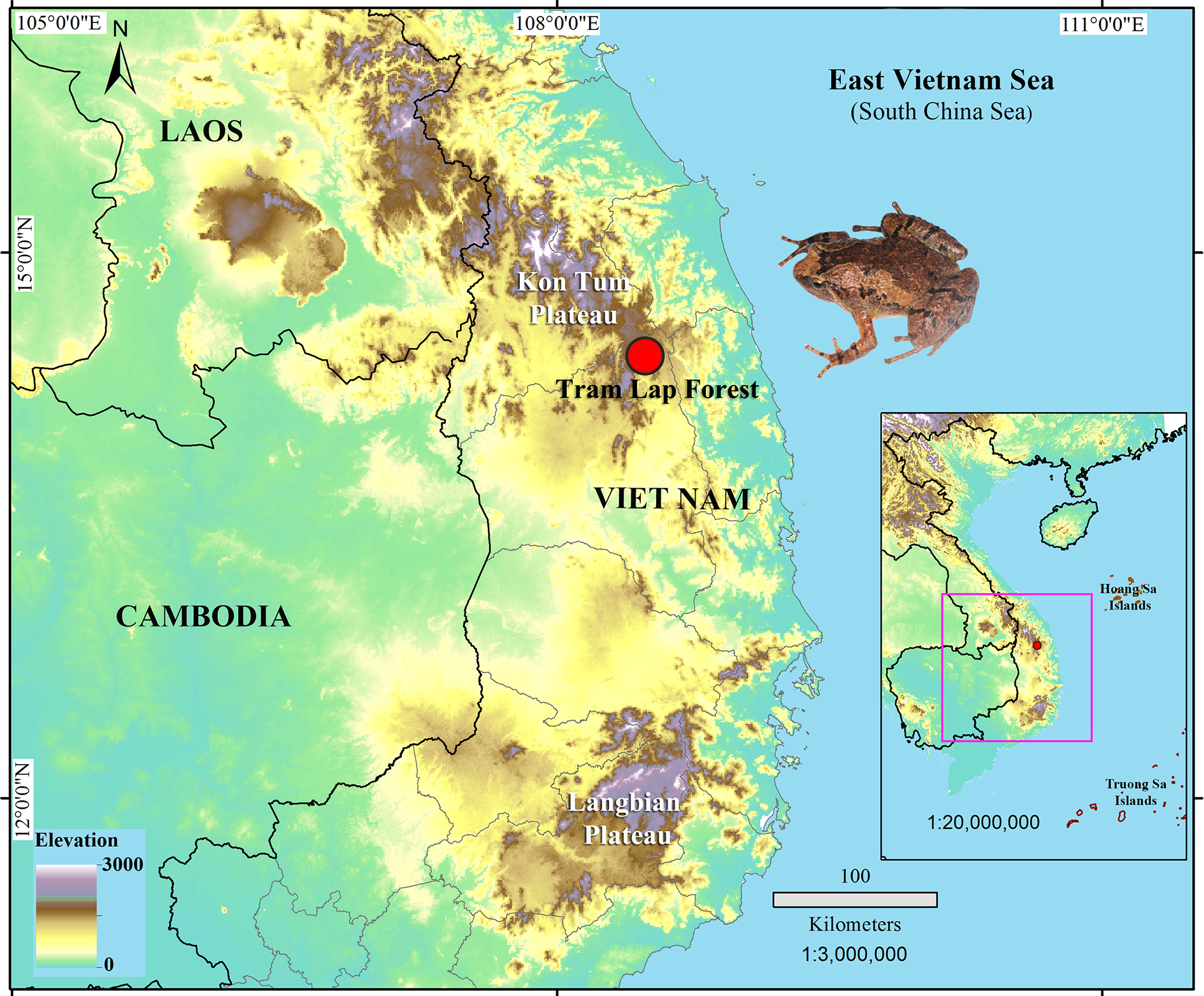
Holotype: ITBCZ 4361 , adult male, collected while calling on the bank of a small pool that formed after heavy rain nearby Ho Chi Minh Tay Road, Tram Lap Forest, Kon Von II Village, Dak Rong Commune , K’Bang District , Gia Lai Province ( 14.53994°N, 108.45104°E, ca. 1210 m a.s.l., Fig. 9 View FIGURE 9 ); collected on 31 July 2016 by Luan T. Nguyen, Tam A. Nguyen, Tiep T. Nguyen, and Vy H. Tran. GoogleMaps
Paratypes: Total 15 specimens, all collected by Luan T. Nguyen, Tam A. Nguyen, Tiep T. Nguyen, and Vy H. Tran at the same location as holotype GoogleMaps : ITBCZ 4288 , adult male, collected on 29 July 2016 ; ITBCZ 4360 , adult gravid female, collected on 0 1 August 2016 ; ITBCZ 4362–4372 , eleven adult males, collected on 31 July 2016 ; ITBCZ 4506–4507 , two adult males, collected on 16 November 2016 .
Diagnosis: Microhyla aurantiventris sp. nov. is characterized by a combination of the following characters: 1) males having SVL 25.2–27.0 mm in 15 individuals, single known female with SVL 30.5 mm; 2) body habitus moderately stocky; 3) head flat, wider than long, snout rounded in dorsal and lateral views, slightly protruding in ventral profile; 4) skin on dorsum and flanks slightly shagreened with numerous tiny tubercles, ventral surfaces smooth; 5) finger I well developed, more than one-half the length of finger II; 6) tips of three outer fingers slightly enlarged, forming weak disks and tips of all toes distinctly dilated into wide disks with narrow peripheral grooves; 7) both finger and toe disks with dorsal median longitudinal grooves; 8) three palmar tubercles (inner and medial palmar tubercles elongated; outer palmar tubercle elongated) and two metatarsal tubercles (inner metatarsal elongated, outer metatarsal rounded); 9) tibiotarsal articulation of adpressed limb reaching slightly beyond the orbit;10) webbing formula: I 1 ¾–2 II 1 ½–2¾ III 2–3 1 / 3 IV 3 ¼–1½ V; 11) dorsum yellowish-brown with dark brown markings, chin and throat yellowish-orange to yellow with small dark brown speckling on its margin, belly bright yellowish-orange to golden-yellow; and 12) an advertisement call consisting of 15–26 pulses with a dominant frequency of 1.1–2.2 kHz (as recorded at 18.5°C). The interspecific genetic p -distances from 12S–16S rRNA gene fragment between the new species and its congeners vary from 9.0% to 14.8%.
Description of holotype ( Figs. 4 View FIGURE 4 , 5 View FIGURE 5 ). Medium-sized specimen, with SVL 26.1 mm; habitus moderately stocky, head length shorter than head width (HL/HW 0.83); snout rounded, slightly prominent in venral profile, longer than diameter of eye (SL/EL 1.25); eye small, slightly protuberant in dorsal view, pupil rounded ( Fig. 5 View FIGURE 5 , F); dorsal surface of head flat, canthus rostralis distinct, rounded; loreal region weekly concave; nostril rounded, laterally, closer to tip of snout than to eye; tympanum hidden, supratympanic fold weak, running from posterior corner of eye to arm insertion; vomerine teeth absent, tongue roundly spatulate and free behind, lacking papillae, vocal sac opening larger.
Forelimbs comparatively short, about three times shorter than hindlimbs (FLL/HLL 0.32); hand longer than lower arm and shorter over two times than forelimb length (HAL/FLL 0.40); fingers slender, rounded in crosssection, first finger well developed, longer by more than one-half of second finger length (1FLO/2FLO 0.64); relative finger lengths: I<IV<II<III (see Fig. 5 View FIGURE 5 , D). Finger webbing and dermal fringes absent; tip of first finger rounded, not enlarged; tips of three outer fingers II–IV slightly dilated forming weak round disks with notable narrow peripheral grooves; relative finger disk widths: II<IV<III; median longitudinal furrow on dorsal surface of fingers II, III, and IV producing appearance of two scutes; subarticular tubercles on fingers distinct, rounded, finger subarticular tubercle formula: 1:1:2:2 (given for fingers I:II:III:IV, respectively); nuptial pad absent; three palmar (metacarpal) tubercles: inner palmar tubercle elongated; outer palmar tubercle almost same length as inner palmar tubercle, elongated in shape (IPTL/OPTL 0.82); medial palmar tubercle elongated and notably bigger than inner and outer palmar tubercles, separated from outer palmar tubercle by a groove; one small rounded supernumerary palmar tubercle present between outer palmar tubercle and first subarticular tubercle of finger IV.
Hindlimbs long and comparatively massive, tibia length slightly longer than half of snout–vent length (TL/ SVL 0.54); tibiotarsal articulation of adpressed limb reaching slightly beyond orbit; foot longer than tibia (FL/TL 1.64); relative toe lengths: I<II<V<III<IV; tarsus smooth, tarsal fold absent on inner surface of tarsus; tips of all toes slightly dilated forming round disks ( Fig. 5 View FIGURE 5 , E), each disk with notable peripheral groove same as on fingers, relative toe disk widths: I<V<II<III<IV; toes slightly flattened, weak dermal fringes present on toes III–IV ( Fig. 5 View FIGURE 5 , E); basal webbing developed between all toes, webbing formula: I 1 ¾–2 II 1 ½–2¾ III 2–3 1 / 3 IV 3 ¼–1½ V; subarticular tubercles on toes distinct, rounded, toe subarticular tubercle formula: 1:2:2:3:2 (for toes I:II:III:IV:V, respectively); nuptial pad absent; two metatarsal tubercles: inner metatarsal tubercle elongated, prominent; outer metatarsal tubercle small, rounded (OMTL/IMTL 0.43).
Dorsal skin slightly shagreened with scattered tiny round tubercles ( Fig. 4 View FIGURE 4 , A, C; Fig. 5 View FIGURE 5 , A, B); few smaller tubercles present on upper eyelids, superciliary projections absent; a weak discontinuous mid-vertebral dermal ridge running from middle of interorbital area towards area above cloaca ( Fig. 5 View FIGURE 5 , A); skin on dorsolateral surfaces smooth; dorsal surface of limbs smooth with few small tubercles, ventral sides of trunk, head, and limbs smooth.
Coloration of holotype in life ( Fig. 5 View FIGURE 5 ). Dorsal surface of head and trunk yellowish-brown; a distinct brown interorbital bar between eyelids, forming a reverse triangle in shape running posteriorly towards scapular region and not covering dorsal surfaces of upper eyelids; a dorsal dark-brown marking consisting of two reverse V-shaped figures forming a hourglass-shaped marking: anterior reverse V-shaped figure runs from scapular area posteriorly and laterally, opening at level of axilla and getting narrower again posteriorly on middle of dorsum; posterior reverse V-shaped figure starts at middle of dorsum and runs laterally towards groin ( Fig. 5 View FIGURE 5 , A, B); two black scapular spots present above axilla on each side of body; few smaller black spots irregularly scattered on dorsolateral and lateral sides of body with one larger dark spot on each side just above axilla; mid-vertebral line brown; supratympanic fold yellowish; dorsolateral surfaces of trunk and upper arm yellowish-brown with a few small black spots; two narrow dark brown bars on each forelimb on dorsal surfaces of forearm; dorsal surfaces of thigh, tibia, and tarsus yellowish-brown with dark brown cross-bars and half cross-bars alternating on each hindlimb: two cross-bars and two half cross-bars on each thigh, one cross-bar and four half cross-bars on each shank, one dark cross-bar on each tarsus ( Fig. 5 View FIGURE 5 , A, B); fingers and toes dorsally beige with dark brown cross-bars; throat and chest dark golden-yellow with dark speckling on margin of throat, ventral skin dark orange-yellow ( Fig. 5 View FIGURE 5 , C), limbs ventrally yellow with small black spots; hand and foot ventrally reddish brown ( Fig. 5 View FIGURE 5 , D, E); pupil black, fine golden reticulations throughout iris except for darker area at anterior corner of iris ( Fig. 5 View FIGURE 5 , F).
Coloration of holotype in preservative. After preservation in ethanol, dorsal coloration changed to light greyish-brown ( Fig. 4 View FIGURE 4 , A, C), ventral surface of chest, belly, and limbs whitish-beige ( Fig. 4 View FIGURE 4 , B); dorsal pattern, dark spots on dorsum and stripes on dorsal surfaces of limbs unchanged, dark brown pattern changed to lighter brown; iris completely black.
Variation ( Table 4, Fig. 6 View FIGURE 6 ). Specimens vary in body size, coloration of dorsal surface, form of dark brown markings on the dorsum, and in the coloration of ventral surfaces: the single gravid female paratype (ITBCZ 4360) has a larger body size than male paratypes (SVL 30.5 mm vs. 26.15± 0.52 mm [25.2–27.0; n=15] in males); males ITBCZ 4365 and ITBCZ 4369 have a much darker dorsal coloration—a dark brown background color with dark blackish-brown markings, male ITBCZ 4365 has less distinct posterior dark brown reverse V-shaped figure on dorsum (see Fig. 6 View FIGURE 6 ); males ITBCZ 4364–4365 and ITBCZ 4368–4369 have more dark spots in the axilla area compared to the holotype. The female paratype ITBCZ 4360 has a lighter yellowish ventral surface (vs. orange to yellowish-orange in males, see Fig. 6 View FIGURE 6 , lower part) and a yellowish-beige throat with brown speckling. Two paratypes (ITBCZ 4506–4507) and four non-vouchered specimens observed on 16 November 2016 have a lighter yellowish ventral coloration compared to males collected in the end of July and in August 2016.
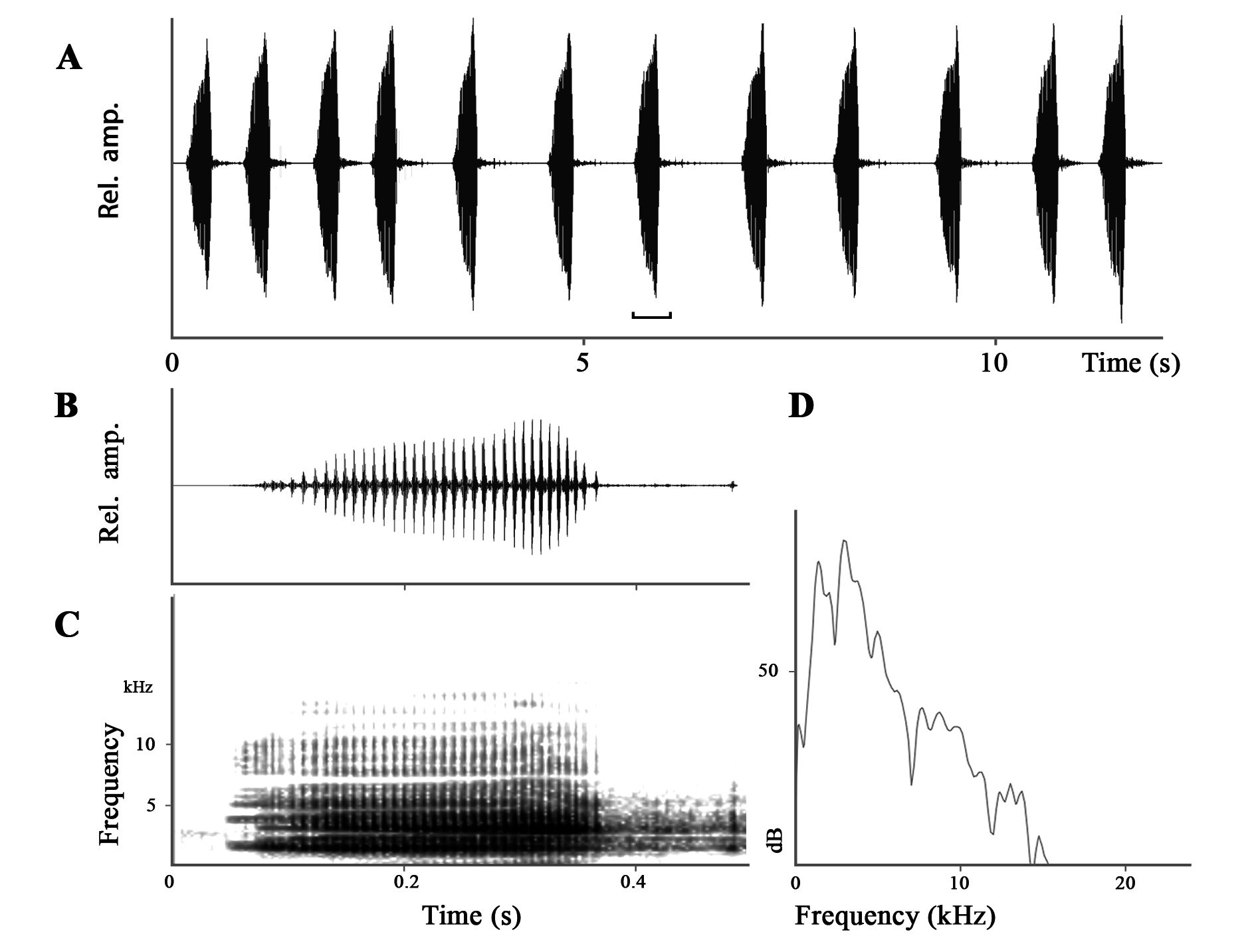
Advertisement call. The advertisement call description is based on recordings of paratypes ITBCZ 5406 and ITBCZ 5407; all calls were recorded at 18.5°C ambient temperature ( Fig. 7 View FIGURE 7 , Table 5). Calls consisted of pulses produced in series, sounding like a quacking duck. Call duration ranged from 110 to 227 ms, and calls were repeated at a rate of 3.1–3.9 calls per second with a 88–391 ms intercall interval. Calls contained from 15 to 26 pulses and had an average pulse rate of 110–144 pulses per second. Dominant frequency varied from 1.8 to 2.2 kHz. Rise-time to reach full amplitude was 67–199 ms, and fall-time from the peak to the end of call varied from 27–48 ms ( Table 5). Harmonics were present at approximately 4.3, 6.3, 10.5, 17.3, and 19.5 kHz. No fundamental frequency was detected ( Fig. 7 View FIGURE 7 ).
Natural history. All specimens were collected at night from 19:00 to 00:00 h on the banks of a small temporary pond that was formed after heavy rain along the sides of a recently constructed road ( Fig. 9 View FIGURE 9 ). Male frogs were found calling from the small dry branch of a dead tree in close proximity to a water-filled pool. The single female was found about 1 m from a temporary pond at night. Until recently, the survey area was covered by evergreen polydominant montane primary forests. Construction of the Ho Chi Minh Road (completed in 2013) across the area damaged forested areas and facilitated formation of temporary waterbodies along the road embankment. Calling males were also heard at around 15:00 h. The new species was found in sympatry with four congeners including M butleri , M. heymonsi , M. mukhlesuri , and M. pulverata ( Fig. 10 View FIGURE 10 ) and microhylids Kaloula indochiensis Chan, Blackburn, Murphy, Stuart, Emmett, Ho & Brown and Micryletta inornata (Boulenger) , all of which were reproducing simultaneously with the new species, sharing the same breeding site. Other anurans such as Feihyla palpebralis (Smith) , Feihyla vittata (Boulenger) , Polypedates sp. ( leucomystax species complex), Rhacophorus kio Ohler & Delorme , R. rhodopus Liu & Hu , Fejervarya limnocharis (Gravenhorst) , and Occidozyga martensii (Peters) also occurred in sympatry. Larval stages and eggs of the new species are unknown.
Distribution. Microhyla aurantiventris sp. nov. is currently known only from the type locality in Tram Lap Forest, Kon Von II Village, Dak Rong Commune, K’Bang District, Gia Lai Province, Vietnam ( Fig. 1 View FIGURE 1 ). The species was recorded from elevation ca. 1210 m a.s.l. The distribution of the new species is unknown, and discovery of new localities within the Kon Tum Plateau is anticipated.
Conservation status. Currently, evergreen forest in Tram Lap connects with other forests in Kon Tum and Gia Lai Provinces. Based on its habitat and distribution elevation, the new species is likely to be endemic to Kon Tum Plateau. However, the extent of its actual distribution range requires further study. Given the available information, we suggest Microhyla aurantiventris sp. nov. be considered as Data Deficient following IUCN’s Red List categories ( IUCN Standards and Petitions Subcommittee 2016).
Etymology. The specific name “ aurantiventris ” is a Latin adjective in the nominative singular, feminine gender, derived from “ aurantiacus ”—“orange-colored” and “ venter ”—“belly”, referring to the distinctive bright orange-yellow coloration of ventral surfaces in adult males of the new species. The recommended common name in English is “Orange-bellied narrow-mouth frog”. The recommended common name in Vietnamese is “Nhái bẫu bụng vàng”.
Comparisons. In having medium body size for Microhyla (SVL 25.2–27.0 mm in 15 males and 30.5 mm in a single female), M. aurantiventris sp. nov. can be distinguished from the smaller members of the genus including M. achatina (male 16 mm, female 23 mm), M. annamensis (males 15.2–19.8 mm, females 18.2–22.6 mm), M. annectens , M. arboricola Poyarkov, Vassilieva, Orlov, Galoyan, Tran, Le, Kretova & Geissler , M. borneensis , M. chakrapanii Pillai , M. fissipes ; M. fusca , M. heymonsi , M. karunaratnei Fernando & Siriwardhane , M. kodial , M. laterite Seshadri, Singal, Priti, Ravikanth, Vidisha, Saurabh, Pratik & Gururaja , M. maculifera Inger , M. malang , M. marmorata , M. minuta Poyarkov, Vassilieva, Orlov, Galoyan, Tran, Le, Kretova & Geissler , M. mixtura , M. mukhlesuri , M. mymensinghensis Hasan, Islam, Kuramoto, Kurabayashi & Sumida , M. nanapollexa , M. nilphamariensis , M. orientalis , M. ornata , M. palmipes , M. perparva , M. petrigena , M. pineticola , M. pulchella , M. pulverata , M. sholigari , M. superciliaris , Microhyla taraiensis , and M. zeylanica ( Table 6).
* The holotype specimen of M. fusca is male (N.A. Poyarkov, unpublished data).
In having moderately stocky body habitus (defined by Bain & Nguyen 2004, Poyarkov et al. 2014), the new species can be distinguished from congeners that have slender or moderately slender bodies, including M. achatina , M. annectens , M. berdmorei , M. butleri , M. fusca , M. mihintalei , M. minuta , M. nanapollexa , M. okinavensis , M. orientalis , M. ornata , M. palmipes , M. sholigari , M. superciliaris , and M. zeylanica .
In having first finger well-developed (1FLO>½2FLO), Microhyla aurantiventris sp. nov. can be diagnosed from M. borneensis , M. nanapollexa , M. palmipes , M. perparva , and M. petrigena , which have first finger nub or bulge. In having outer metatarsal tubercle oval-shaped, M. aurantiventris sp. nov. can be separated from the burrowing species M. mihintalei , M. picta Schenkel , and M. rubra , which have enlarged shovel-shaped outer metatarsal tubercles used for digging.
In having comparatively short hindlimbs (tibiotarsal articulation of adpressed hindlimb reaching slightly beyond the orbit but not reaching the tip of snout) the new species can be distinguished from M. achatina , M. annamensis , M. annectens , M. arboricola , M. darevskii , M. fissipes , M. mantheyi , M. marmorata , M. mukhlesuri , M. mymensinghensis , M. nanapollexa , M. perparva , M. petrigena , M. pulchella , M. pulchra , M. pulverata , and M. superciliaries , which have tibiotarsal articulation of an adpressed hindlimb reaching just, or well, beyond the snout.
In having weak disks on fingers II–IV, well-developed disks on all toes, and median longitudinal grooves on dorsal surfaces of fingers II–IV and all toes, M. aurantiventris sp. nov. differs from the M. ornata species group ( M. ornata , M. rubra , M. mihintalei , M. nilphamariensis , and M. taraiensis ) and the M. fissipes species group ( M. fissipes , M. mukhlesuri , M. mymensinghensis , and M. okinavensis ), as well as from M. picta , M. pulchra , and M. zeylanica ; these species do not have disks on fingers and toes and lack median longitudinal grooves on dorsal surface of digits. The new species can be diagnosed from M. chakrapanii , M. darevskii , and M. maculifera , which have no disks on fingers and lack median longitudinal grooves on dorsal surface of digits. Microhyla aurantiventris sp. nov. can be further differentiated from M. kodial , M. mixtura , M. sholigari , and M. superciliaries , which have no dorsal median longitudinal grooves on finger disks.
The new species differs from M. annectens , M. arboricola , M. maculifera , M. perparva , and M. petrigena of M. annectens species group by having two metatarsal tubercles (vs. one metatarsal tubercle).
In having toes with basal webbing (webbing formula I 1 ¾–2 II 1 ½–2¾ III 2–3 1 / 3 IV 3 ¼–1½ V), M. aurantiventris sp. nov. can be distinguished from the following members of its genus that have fully developed webbing reaching to disks at most toes (usually with the exception of toe IV): M. annamensis , M. annectens , M. berdmorei , M. darevskii , M. malang , M. mantheyi , M. marmorata , M. nanapollexa , M. perparva , M. petrigena , M. pulchella , M. pulverata , and M. superciliaris (detailed by Poyarkov et al. 2014). Toe webbing of M. arboricola and M. pulchra reaches the level of distal subarticular tubercles and, thus, they can be diagnosed from the new species. The following species of Microhyla show less developed rudimentary toe-webbing not exceeding the level of proximal subarticular tubercles: M. achatina , M. borneensis , M. fissipes , M. heymonsi , M. karunaratnei , M. minuta , M. mixtura , M. mukhlesuri , M. mymensinghensis , M. okinavensis , M. orientalis , M. ornata , M. palmipes , M. picta , M. pineticola , M. rubra , M. sholigari , and M. zeylanica ( Poyarkov et al. 2014) .
By possessing a weak, discontinuous dorsomedial (vertebral) line, M. aurantiventris sp. nov. can be distinguished from M. achatina , M. heymonsi , M. nilphamariensi s, and M. pineticola , which have a distinct and continuous light dorsomedial line. The new species can be further separated from M. heymonsi and M. pineticola by absence of characteristic ()-shaped black spot in the middle of dorsum (vs. present in these species) and by light coloration of head and body flanks (vs. darker coloration of lateral sides of head and body in M. heymonsi and M. pineticola ). The new species can be further distinguished from M. superciliaris by lacking (vs. having) superciliary spines.
By males having in life yellowish-orange to dark golden-yellow throat and vocal sacs with black speckling on its margins, the new species can be distinguished from all known congeners for which coloration of throat and vocal sac in life is described. Microhyla chakrapanii , M. maculifera , M. nanapollexa , M. perparva , and M. sholigari are known from preserved specimens only and coloration in life has not been described. Notwithstanding, in preservative these species have dark to brown or grey coloration on their throats with black or dark mottling or speckling. By having in life yellowish-orange to yellow ventral coloration in reproductively active males, M. aurantiventris sp. nov. differs from all other species of Microhyla for which the ventral coloration in life is described, except for M. berdmorei and M. pulchra , which have lemon-yellow marking in posterior part of belly and groin. In contrast, M. annamensis , M annectens , M. arboricola , M. borneensis , M. maculifera . M. marmorata , M. mihintalei , M. petrigena , M. pulchella , and M. pulverata have dark brown, brownish or grey belly with or without dusty whitish marbling or mottling, M. darevskii , M. fissipes , M. heymonsi , M. laterite , M. malang , M. mantheyi , M. minuta , M. mixtura , M. mukhlesuri , M. mymensinghensis . M. nilphamariensis , M. okinavensis , M. orientalis , M. ornata , M. palmipes , M. picta , M. pineticola , M. rubra , M. superciliaris , M. taraiensis , and M. zeylanica have white to whitish or cream ventral coloration with or without grey/black mottling, and M. karunaratnei has ventral marbling in black and white.
The advertisement call of M. aurantiventris sp. nov. is unique among congeners for which calls have been described. Compared to most of other species, the new species has a relatively low dominant frequency (1.8–2.2 kHz, 18.5 o C), which differs from calls of M. borneensis (3.4–3.7 kHz, 21.7 o C), M. karunaratnei (3.1–3.4 kHz, 19.1 o C), M. korial (3.3–4.2 kHz, 25.2 ± 0.24°C), M. laterite (3.5–3.6 kHz, 27.9 o C), M. marmorata (2.7–3.0 kHz, 17.5 o C), M. petrigena (3.8–5.1 kHz, 24.1 o C), M. sholigari (3.5–3.7 kHz, 23.2 o C), and M. taraiensis (3.1–3.4 kHz, 28.6 o C). In having comparatively high pulse rate (110–144 pulses/s), the call of the new species differs from those of M. berdmorei (33–35 pulses/s), M. borneensis (39–42 pulses/s), M. mukhlesuri (53–60 pulses/s; data for a Thai population recorded previously as M. fissipes ; see Yuan et al. 2016), M. heymonsi (23–39 pulses/s), M. karunaratnei (65–87 pulses/s), M. malang (27.7–32.2 pulses/s), M. mihintalei (50–66 pulses/s), M. nilphamariensis (35.4–36.5 pulses/s), M. ornata (37–45 pulses/s), M. petrigena (89 pulses/s), M. rubra (108 pulses/s), and M. zeylanica (37–49 pulses/s).
In having 15–26 pulses per call, the call of M. aurantiventris sp. nov. differs from calls of species having a higher number of pulses per call (50–95 pulse/call in M. karunaratnei , 90–126 pulses/call in M. laterite , 38–80 pulses/call in M. pulchra , 64–72 pulses/call in M. sholigari , and 61–92 pulses/call in M. zeylanica ) and from congeners with lower number of pulses per call (2–4 pulses/call in M. borneensis , 2–7 pulses/call in M. korial , 4–8 pulses/call in M. malang , 9–14 pulses/call in M. ornata ). The call duration of the new species ranges between 110 ms and 227 ms which differs from those of M. mukhlesuri (230–310 ms; data for population from Thailand recorded previously as M. fissipes [ Yuan et al. 2016]), M. karunaratnei (699–1172 ms), M. laterite (600–850 ms), M. marmorata (554–1443 ms), M. nilphamariensis (mean 410 ms), M. ornata (mean 280 ms), M. sholigari (530– 810 ms), and M. zeylanica ( 1503–1999 ms).
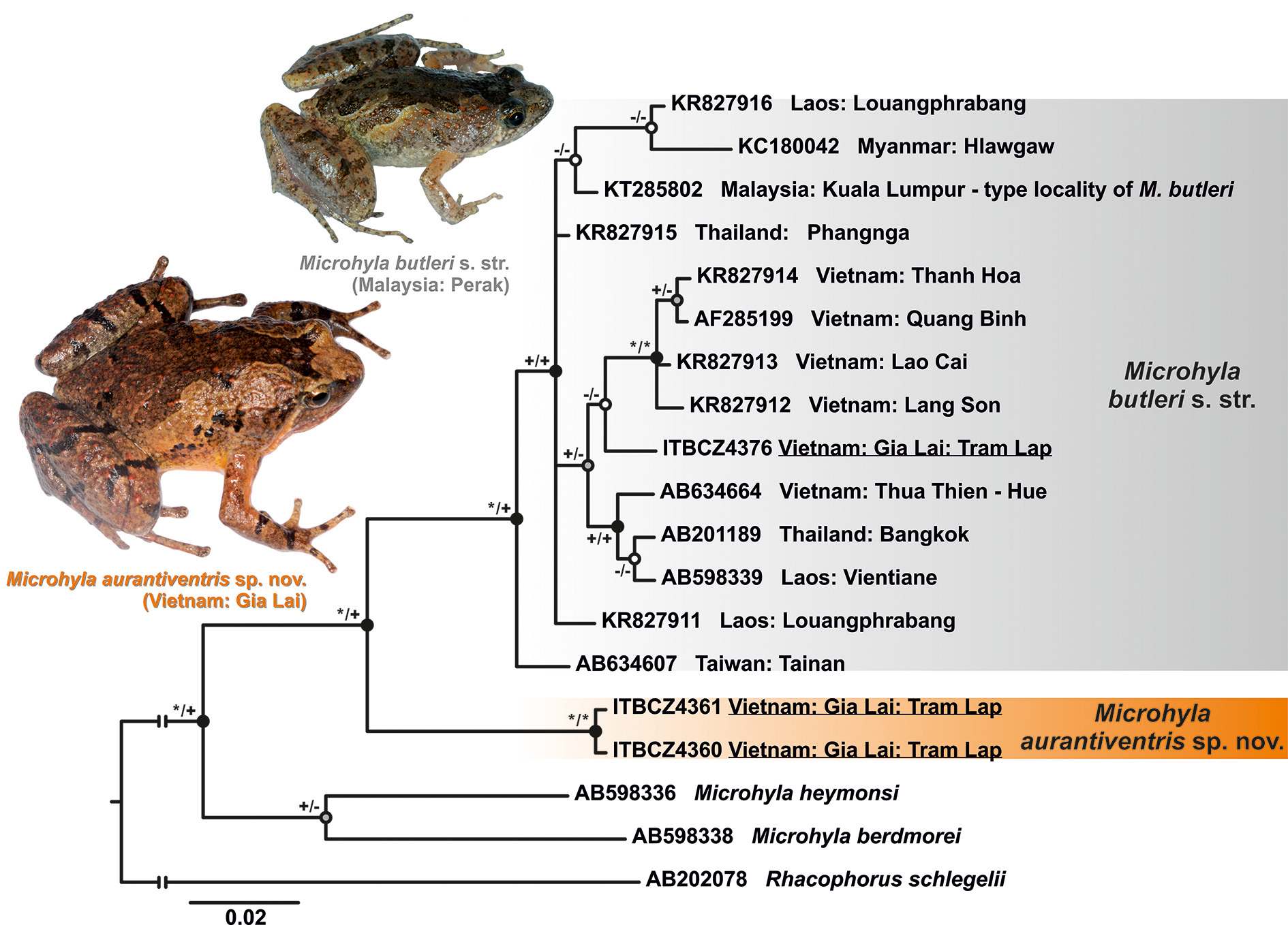
Superficially, Microhyla aurantiventris sp. nov. most closely resembles its sympatric sister-species M. butleri , but several morphological characters distinguish them. Microhyla aurantiventris sp. nov. is larger than M. butleri , with male SVL 25.2–27.0 mm, and 30.5 mm in the single known female (vs. adult SVL in males 20.0–25.0 mm, and in females 21.0–26.0 mm of M. butleri ; Table 6); values for known adult SVL ranges do not overlap for the two species. The holotype of M. butleri (female, according to Parker 1934) has a SVL of 21 mm ( Boulenger 1900). Male M. butleri collected at the type locality of the new species have SVL values of 20.6–21.0 mm (n=5) and a single female had a SVL of 23.9 mm, thus corresponding well to the body size of M. butleri sensu stricto. Males of M. butleri have SVL values of 20.1–25.0 mm and females have have SVL values of 19.4–26.0 mm ( Poyarkov et al. 2014; Taylor 1962; Ziegler 2002 [ 1 female, 3 males], from Ha Tinh Province, one specimen [TZ 9874, GenBank: AF285199 View Materials ] used in our molecular analysis; and our collection from Khanh Hoa Province, Vietnam [ 2 females, 6 males]). In addition, Taylor (1962) provided data for one specimen from Kuala Tahan, Malaysia (EHT-HMS No. M. 162, SVL 26 mm) and the SVL 26 mm was also mentioned by Bourret (1942) and Parker (1934) without sex provided. These specimens were not used to compare with the new species due to their uknown sex. The new species also has more extensive webbing on toes than M. butleri (webbing formula I 1 ¾–2 II 1 ½–2¾ III 2–3 1 / 3 IV 3 ¼–1½ V in the new species vs. I 2–2 ½ II 1 ¾–3 III 2 1 / 3 –3½ IV 3 ½–2¼ V in sympatric M. butleri , our data).
The new species also has a notably more stocky body habitus than M. butleri ( Fig. 3 View FIGURE 3 ), which is reflected in a number of diagnostic morphometric character ratios to SVL (below as mean ± SD and ranges for 11 males of the new species and three sympatric males of M. butleri s. stricto; statistical tests were not implemented due to small sample size for M. butleri s. stricto). Microhyla aurantiventris sp. nov. generally has shorter head than M. butleri with HL/HW 0.64–0.88 (0.79±0.07) (vs. 0.82–0.95, 0.87±0.06, in M. butleri ) and HL/SVL 0.26–0.34 (0.31±0.03) (vs. 0.32–0.38; 0.36±0.03, in M. butleri ). Microhyla aurantiventris sp. nov. has also shorter forelimbs and generally shorter hindlimbs than M. butleri : FLL/SVL 0.56–0.64 (0.60±0.03) (vs. 0.67–0.74, 0.72±0.03) and HLL/ SVL 1.70–2.07 (1.85±0.10) (vs. 1.89–1.97, 1.95±0.04).
The coloration of Microhyla aurantiventris sp. nov. in life also distinguishes it from M. butleri ( Fig. 3 View FIGURE 3 vs. Fig. 10 A View FIGURE 10 ). Microhyla butleri always lack the characteristic orange-yellow coloration of belly and throat found in males of the new species as well as the light middorsal line; breeding males have whitish to greyish-beige venters, throat with dense black speckles and no light middorsal line. Further, M. butleri have a dark-brown hourglass-shaped figure on dorsum edged with white to beige line, and dorsal tubercles in breeding males have reddish to orange tint. In contrast, male M. aurantiventris sp. nov. do not have light edging on the dorsal hourglass-shaped dark figure and reddish or orange tubercles (our data; Manthey and Denzer, 2014).
The advertisement call of M. aurantiventris sp. nov. differs from calls of its sister-species M. butleri recorded in sympatry ( Fig. 8 View FIGURE 8 ) by having lower dominant frequency (1.8–2.2 kHz, 18.5 o C vs. 2.9–3.0 kHz, 18.0 o C in M. butleri ), shorter intercall interval (88–391 ms, mean 154.5 ms vs. 357–2034 ms, mean 837.8 ms in M. butleri ), shorter call rise time (67–199 ms vs. 224–920 ms in M. butleri ), shorter call duration (110–227 ms vs. 301–354 ms in M. butleri ), and lower number of pulses per call (15–26 pulses/call vs. 34–40 pulses/call in M. butleri ). Although frequency can vary with temperature, differences of the advertisement call between two sympatric species on the scale reported here are very unlikely to be attributed to temperature (0.5 o C in temperature differential).
Finally, the new species is markedly distinct from all other congeners for which comparable sequences are available, including its closest relative M. butleri , by relatively large uncorrected genetic distances ( p ḵ9.0%) at the 12S rRNA–16S rRNA mtDNA gene fragment examined.






No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.