
Enicospilus myricae, Broad & Shaw, 2016
|
publication ID |
https://doi.org/10.5852/ejt.2016.187 |
|
publication LSID |
lsid:zoobank.org:pub:8ACE88A9-6CC8-4824-837B-3F20311E7957 |
|
DOI |
https://doi.org/10.5281/zenodo.3852440 |
|
persistent identifier |
https://treatment.plazi.org/id/31771CB3-729B-4157-A201-B6736BBB2800 |
|
taxon LSID |
lsid:zoobank.org:act:31771CB3-729B-4157-A201-B6736BBB2800 |
|
treatment provided by |
Valdenar |
|
scientific name |
Enicospilus myricae |
| status |
sp. nov. |
Enicospilus myricae sp. nov.
urn:lsid:zoobank.org:act:
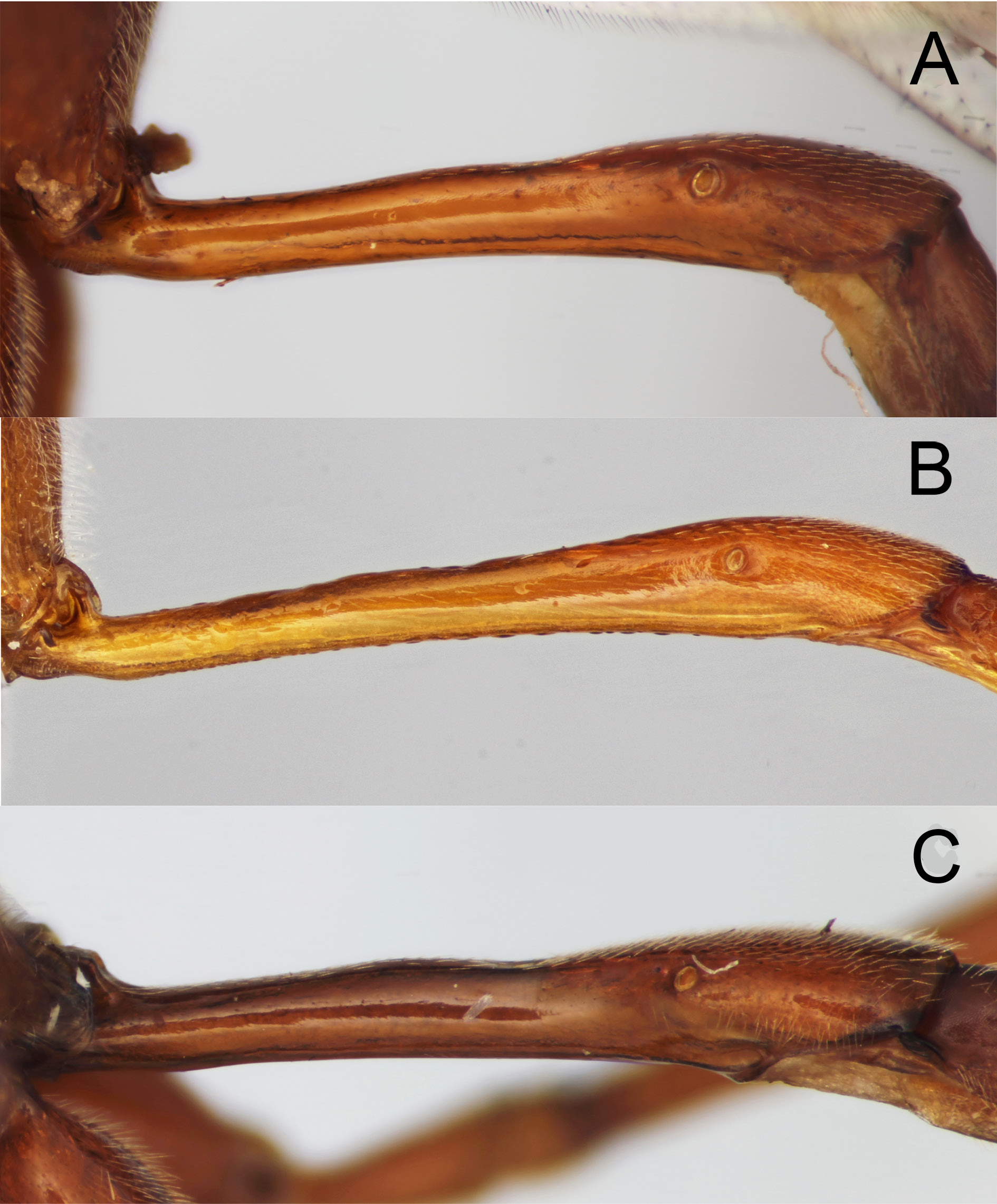
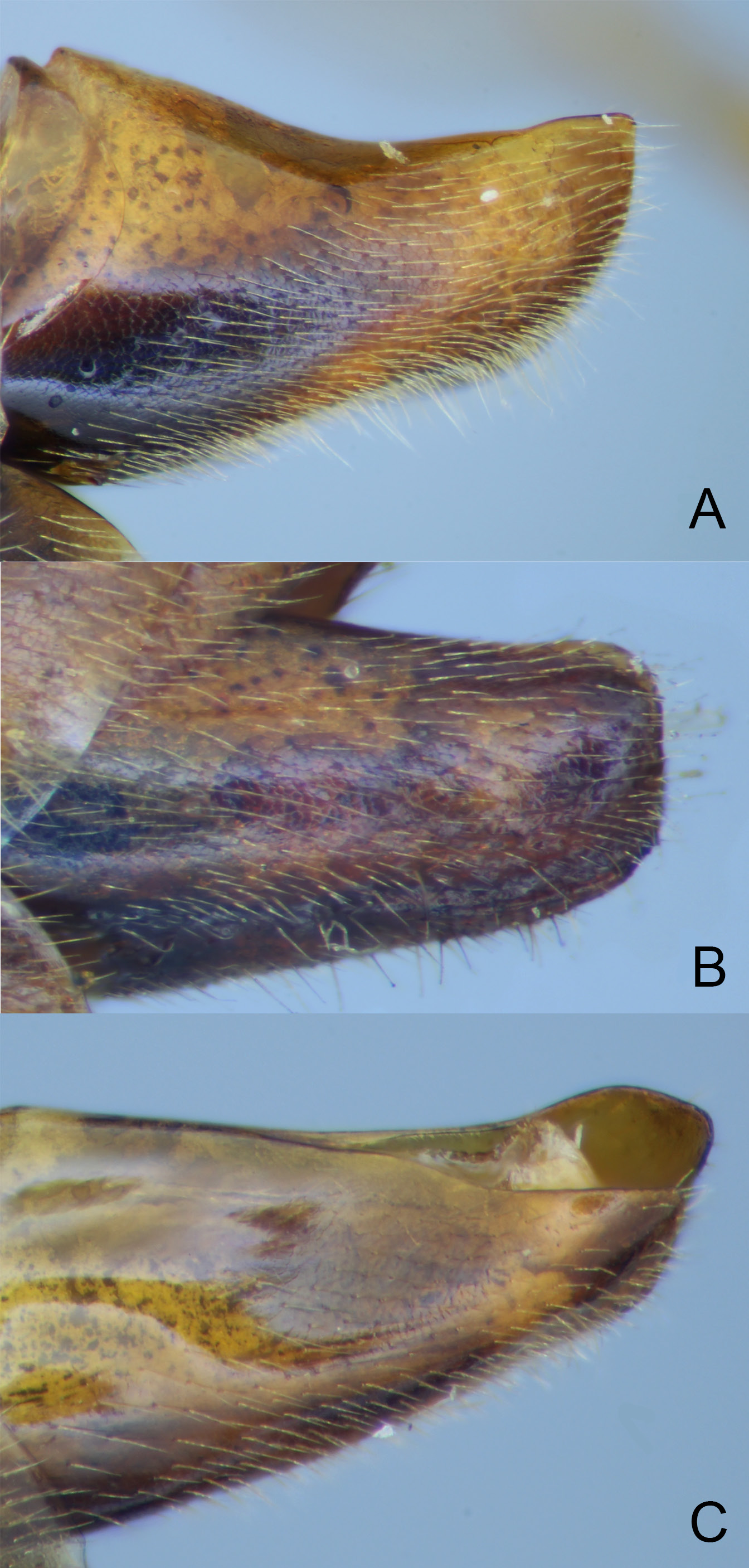
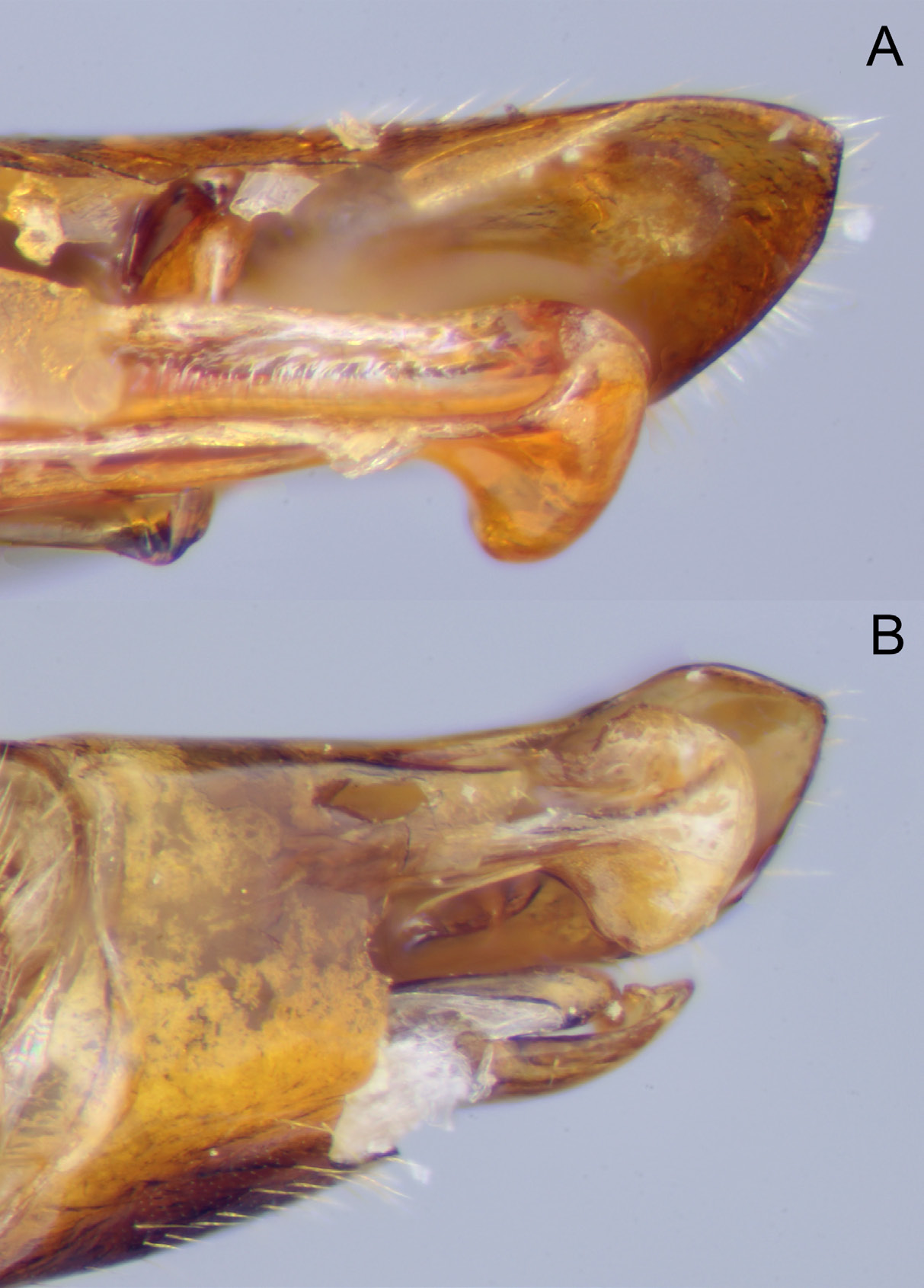
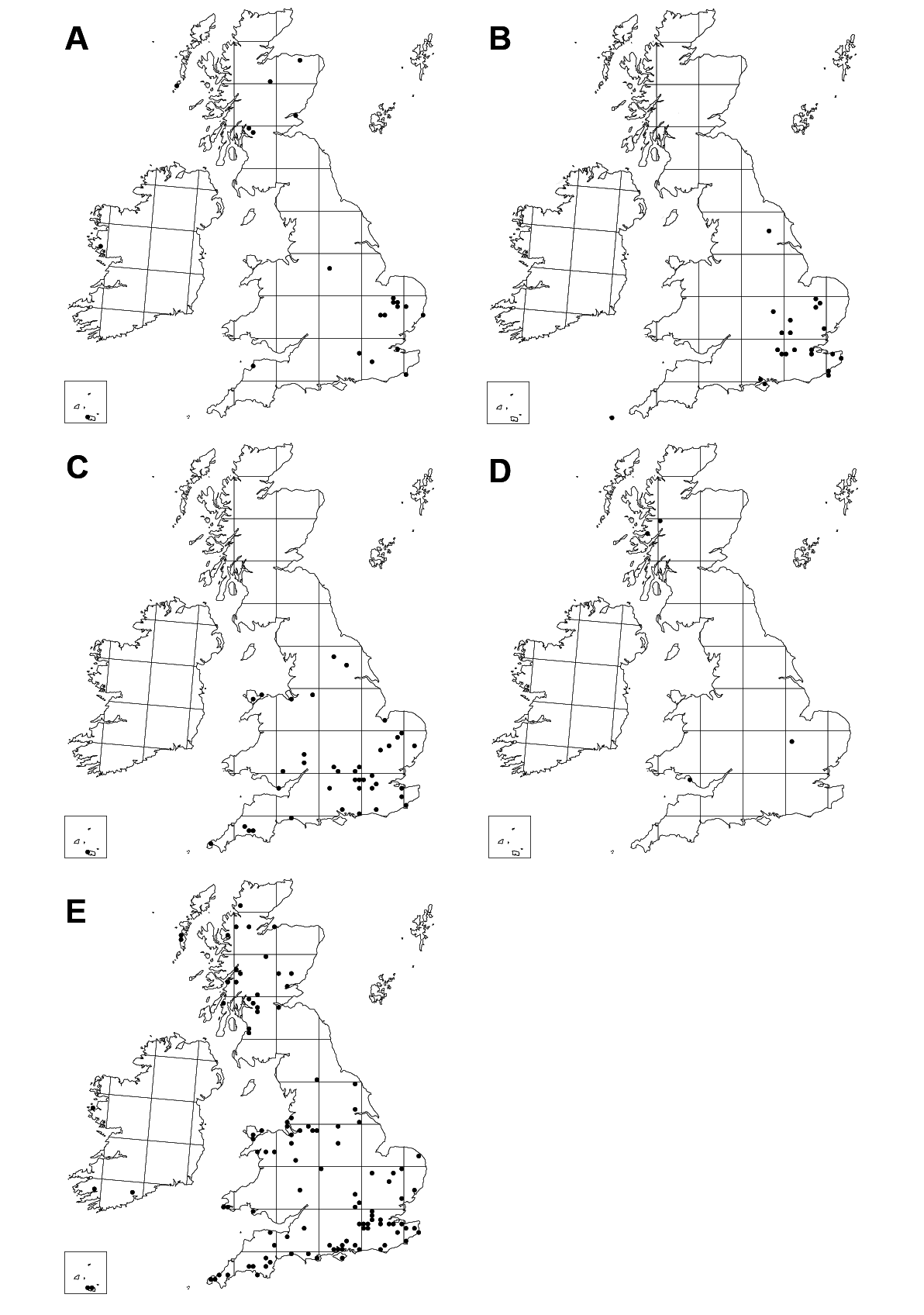
Figs 4C View Fig , 5C View Fig , 6C View Fig , 8C View Fig , 9B View Fig , 10 View Fig , 16 View Fig , 18D View Fig
Diagnosis
Separated from E. adustus and E. cerebrator by the more rounded temples, wider ocellar-ocular space (especially compared to E. cerebrator ), more obvious dorsal “dip” on the first tergite and by the distinctly different aedeagus of the male.
Etymology
Named after the association with the distinctive habitat of Myrica gale -dominated bog, from which this species was reared.
Material examined
Holotype
SCOTLAND: ♀: “[Scotland] Resipole, Argylls. NM 723645. Orthosia gracilis , Myrica gale coll. 4.7.92, PLE [parasitoid larva erupted] 7.92 em. 7.1993 M. R. Shaw” ( NMS).
Paratypes
SCOTLAND: 2 ♀♀, same data as holotype ( NMS); 1 Ƌ, Resipole, Argyllshire, coll. 4 Jul. 1992 (M. R. Shaw) ( NMS); 1 ♀, 5 ƋƋ, Loch Arkaig, Invernesshire NN0291, Malaise trap in native pinewood, Jun. 1992 (I. MacGowan) ( NMS).
WALES: 1 ♀, Glamorgan, Kenfig, 30 Jun. 1963 ( R.B. Benson) ( BMNH). ENGLAND: 1 ♀, Huntingdonshire, Monks Wood NNR, 17–29 Jul. 1978 (M.G. Fitton & J.S. Noyes) ( BMNH).
AUSTRIA: 1 ♀, Burgenland, Leitha Gebirge, 10–23 Jun. 1956 (E.W. & I.D. Classey) ( BMNH).
Description
Female
There is very little variation between specimens; variation is covered in the description. Fore wing length 11–13.5 mm. Antenna with 57–59 flagellar segments (n = 6) ( 57 in holotype). 1 st flagellomere about 5× as long as apically wide, preapical flagellomere 1.5× as long as wide. Head ( Fig. 4C View Fig ) with distinct gap between lateral edge of stemmaticum and edge of eye, lateral ocellus separated from eye by 0.25× maximum length of ocellus; temples in dorsal view curved, rounded immediately behind eye, then more linearly narrowed, measured in straight line from eye margin to lateral margin of occipital carina, c. 0.8× length of greatest eye width; in lateral view, at level of antennal sockets, gena 0.8× width of eye. Mandible strongly bent, slightly twisted, with curved groove containing long setae, from dorsal proximal corner to base of teeth, as in other members of E. ramidulus group; lower tooth about 0.5× length of upper. Clypeus apically truncate with wide, thin flange, very sparsely punctate. Eyes ventrally slightly convergent, face at mid-height 1.6× as wide as high, evenly, fairly closely punctate. Mesopleuron entirely, closely punctate, with faint transverse striae across dorsal portion below subalar prominence and more prominently in narrow band along dorsal 0.5 of posterior edge (deflected anteriorly by unsculptured speculum) and across wide area medio-ventrally on mesopleuron; Austrian paratype with striation reduced, only noticeable on medio-ventral area of mesopleuron. Epicnemial carina almost complete, fading out dorsally just before reaching anterior edge of mesopleuron. Mesoscutum with notauli faintly indicated anteriorly, entirely closely punctate (punctures small, closer than puncture diameter), shiny. Scutellum ( Fig. 5C View Fig ) shiny, regularly punctate, punctures larger than on mesoscutum and further apart (larger and closer anteriorly); lateral carinae complete to near posterior end of scutellum, indicated around posterior end of scutellum as rugosity/carinulae. Fore wing ( Fig. 10 View Fig ) as in other species of E. ramidulus group; glabrous area (fenestra) of discosubmarginal cell extending from proximal sclerite to posterior 0.3 of Rs+ 2 r (along thickened area of vein); proximal sclerite entirely pigmented, approximately triangular with rounded anterior angle, more elongate on distal corner; central sclerite roughly “D”-shaped, pigmented distally, fading to unpigmented, transparent proximally; distal sclerite represented by faintly pigmented line along distal-ventral margin of fenestra; fore wing vein Rs+ 2 r sinuous , uniformly widened along anterior 0.7; 3 rs-m 0.45× section of M between 1 m-cu and 3 rs-m; cu-a slightly to distinctly proximal to Rs&M. Metapleuron shiny, closely punctate. Propodeum with weakly defined central section of anterior transverse carina, anterior of this shiny and superficially punctate, posterior to this entirely reticulate-rugose or sculpture much reduced dorso-laterally. First metasomal tergite ( Fig. 6C View Fig ) with shallow dorsal concavity at anterior 0.45. Second metasomal segment with laterotergite narrow, folded under; third tergite with laterotergite not demarked.
Colour
Uniformly testaceous ( Fig. 16 View Fig ), varying from dull orange to a darker, reddish-orange (although probably dependent on preservation), except for black mandibular teeth and varying amounts of dark brown infuscation on the venter of the metasoma from 4th tergite onwards, and apical tergites at most weakly infuscate. Antenna darker apically. Generally slightly darker than E. adustus .
Male
As in female but with more antennal segments (61–64 flagellar segments; n = 4) and striation on mesopleuron much feebler, basically absent medio-ventrally. Paramere ( Fig. 8C View Fig ) rather strongly narrowed posteriorly, smoothly angled into apical edge; aedeagus ( Fig. 9B View Fig ) with dorsal, apical area concave and laterally carinate, more rounded apically than in E. adustus ( Fig. 9A View Fig ) or E. cerebrator , in which apex of aedeagus less protruding dorsally and more strongly reflexed ventrally.
Distribution
Austria, England, Scotland, Wales, as detailed in the list of type material (British distribution in Fig. 18D View Fig ).
Flight time (non-reared material)
June-July.
Hosts
Orthosia gracilis (Denis & Schiffermüller, 1775) ( 3 specimens, from one collecting event) ( Noctuidae : Hadeninae ).
The majority of the few known specimens were collected in Scotland but it is a much more widespread species and it may be that it prefers boggy habitats in which few people collect ichneumonids. One paratype was collected in Monks Wood NNR, an ancient deciduous woodland with a rather rich fauna of fen or bog-associated noctuids. Unlike other British Enicospilus there is a distinct sexual dimorphism in antenna length, as males have more flagellar segments, with no overlap in the small sample size available.




| R |
Departamento de Geologia, Universidad de Chile |
| NMS |
National Museum of Scotland - Natural Sciences |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SuperFamily |
Ichneumonoidea |
|
Family |
|
|
SubFamily |
Ophioninae |
|
Genus |