
Proceratophrys Ʋelhochico, 2022
|
publication ID |
https://doi.org/ 10.1670/20-070 |
|
DOI |
https://doi.org/10.5281/zenodo.7887606 |
|
persistent identifier |
https://treatment.plazi.org/id/039187F4-FFC4-FFAE-D8DE-F8C1AC7BB0C8 |
|
treatment provided by |
Diego |
|
scientific name |
Proceratophrys Ʋelhochico |
| status |
sp. nov. |
Proceratophrys Ʋelhochico View in CoL View at ENA sp. nov.
( Figs. 2–4 View FIG View FIG View FIG , Fig. 8 View FIG ; Tables 3–5 View TABLE View TABLE View TABLE )
Holotype.— ZUFMS-AMP13650 (adult male), Delfino district, Campo Formoso municipality, Bahia state (10°19, 2.27"S, 41°9, 11.62"W; 850 m a.s.l.), collected on 19 March 2018 by F.M. Magalhães and W. Pessoa.
124 Paratopotypes.— UFMG 6224–6225 View Materials (adult females), UFMG 6220 View Materials (juvenile) collected in the type locality on 16 December 2010 by F. Leite, M. Lindermann and C. Rievers .
Paratypes.— Total of 12 individuals. ZUFMS-AMP13652 (adult male) collected in Umburanas municipality, Bahia State (10°27, 52 ,, S, 41°23, 26 ,, W; 1,040 m a.s.l., datum = WGS84 ), on 30 August 2019 by D. Cavalheri. ZUFMS-AMP 13942 , 13943 , 13945 , 13947 , 13948 (adult males), ZUFMS-AMP 13944 , 13946 (adult females), ZUFMS-AMP 13950 (juvenile) collected in Umburanas municipality, Bahia State, on 4 November 2020 by D. Cavalheri. ZUFMS-AMP 13949 (adult female) collected in Umburanas municipality, Bahia State, on 8 November 2020 by D. Cavalheri. ZUFMS-AMP 14724 (adult male), ZUFMS-AMP 14725 (adult female) collected in Umburanas municipality, Bahia State, on 8 February 2021 by D. Cavalheri.
126
Diagnosis.—Diagnosed by the following combination of characters: 1) small to medium size (32.1–38.7 mm in adult males, 37.7–44.3 mm in adult females; Table 3 View TABLE ); 2) snout rounded in dorsal and ventral views, vertical in profile; 3) small and rounded tubercle at the buccal commissure; 4) upper eyelid border wrinkled (absence of well-defined tubercles like those found on the palpebral region); 5) dorsal skin with few sparse and short warts (inter-ocular and ocular-dorsal ridge of warts absent); 6) one row of small, rounded, and not fused tubercles on the forearm; 7) presence of a single rounded prominent calcar tubercle; 8) ventral region cream without spots; 9) advertisement call with a single multipulsed note of 29–39 pulses, lasting 0.327 – 0.442 sec, and dominant frequency of 1313 Hz.
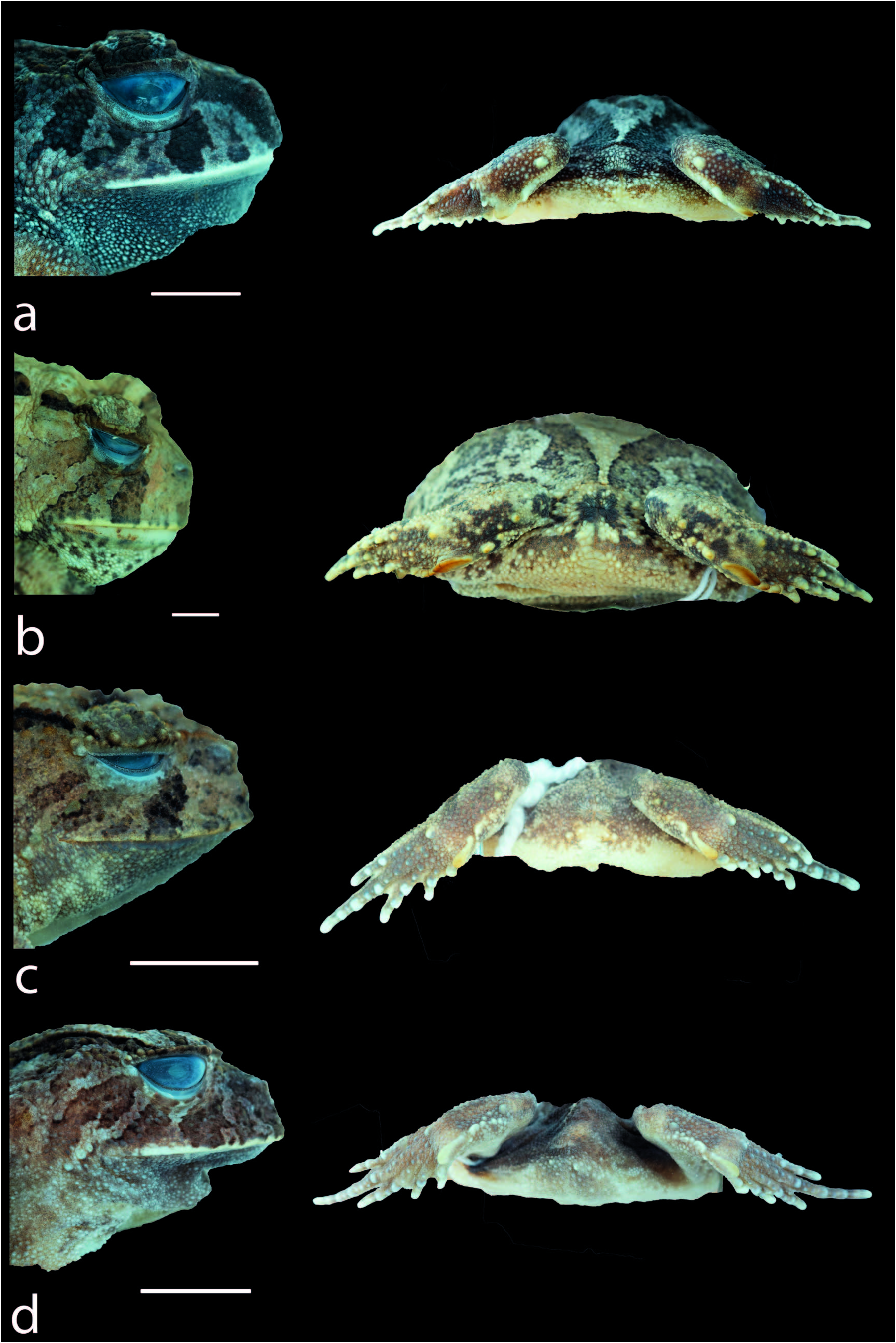
Comparisons aeith Other Species.— Proceratophrys oelhochico sp. nov. is readily diagnosed from all species of Proceratophrys by the presence of a small and rounded tubercle on the buccal commissure ( Figs. 3 View FIG , 4c View FIG ; enlarged and generally triangular/ pointed in other Proceratophrys , small in some individuals of P. cristiceps ; Fig. 5 View FIG ), by the upper eyelid border wrinkled with tubercles indistinct from the others of the dorsal region ( Fig. 3a View FIG ) (presence of single uni-cuspidate palpebral appendage or a large marginal row of tubercles on the eyelid in other Proceratophrys ; Fig. 5 View FIG ), by the presence of few sparse and short warts on the dorsal skin, with no ocular-dorsal ridge of warts ( Figs. 2a View FIG , 4 View FIG ; dorsum scattered with several small, pointed and/or rounded granules in other Proceratophrys , few sparse and short warts in one individual of P. redacta ; Fig. 5 View FIG ), and presence of a single rounded prominent calcar tubercle ( Figs. 3a View FIG , 4e View FIG ; tarsus covered with warts and tubercles in various sizes in other Proceratophrys ; Fig. 3 View FIG ).
Proceratophrys oelhochico sp. nov. can also be distinguished from P. appendiculata , P. belzebul , P. gladius , P. itamari , P. izecksohni , P. laticeps , P. mantiqueira , P. melanopogon , P. moheringi , P. phyllostomus , P. pombali , P. sanctaritae , P. subguttata , and P. tupinamba by lacking a rostral appendage (present in those species). The absence of postocular swellings distinguish Proceratophrys oelhochico sp. nov. from P. aoelinoi , P. bigibbosa , P. brauni , and P. palustris (present). The absence of a complete ocular-dorsal ridge of warts distinguishes Proceratophrys oelhochico sp. nov. from P. bagnoi , P. goyana , P. redacta , and P. rotundipalpebra (complete ocular-dorsal ridge of warts). From P. bagnoi , P. concaoitympanum , P. dibernardoi , P. goyana , and P. huntingtoni , Proceratophrys oelhochico sp. nov. differs by the presence of only one row of tubercles in the forearm (two rows in P. bagnoi , P. concaoitypanum , P. cristiceps , P. dibernardoi , and P. huntingtoni , tubercles not organized in rows in P. goyana ). The presence of ventral region cream, without spots or vermiculations, distinguishes Proceratophrys oelhochico sp. nov. from P. ararype , P. branti , P. carranca , P. huntingtoni , P. redacta , and P. strussmannae (dark-brown mottling on the gular region, chest and belly in P. ararype , dark spots in P. branti , P. carranca , and P. huntingtoni , gular region blackish, scattered small, dark-brown dots in the chest in P. redacta , and in the chest and belly in P. strussmannae ). By the presence of only one multipulsed note in the advertisement call, Proceratophrys oelhochico sp. nov. is also distinguished from P. carranca (1–10 notes/call), P. goyana (1–34), P. rotundipalpebra (1–24), and P. oielliardi (3–20).
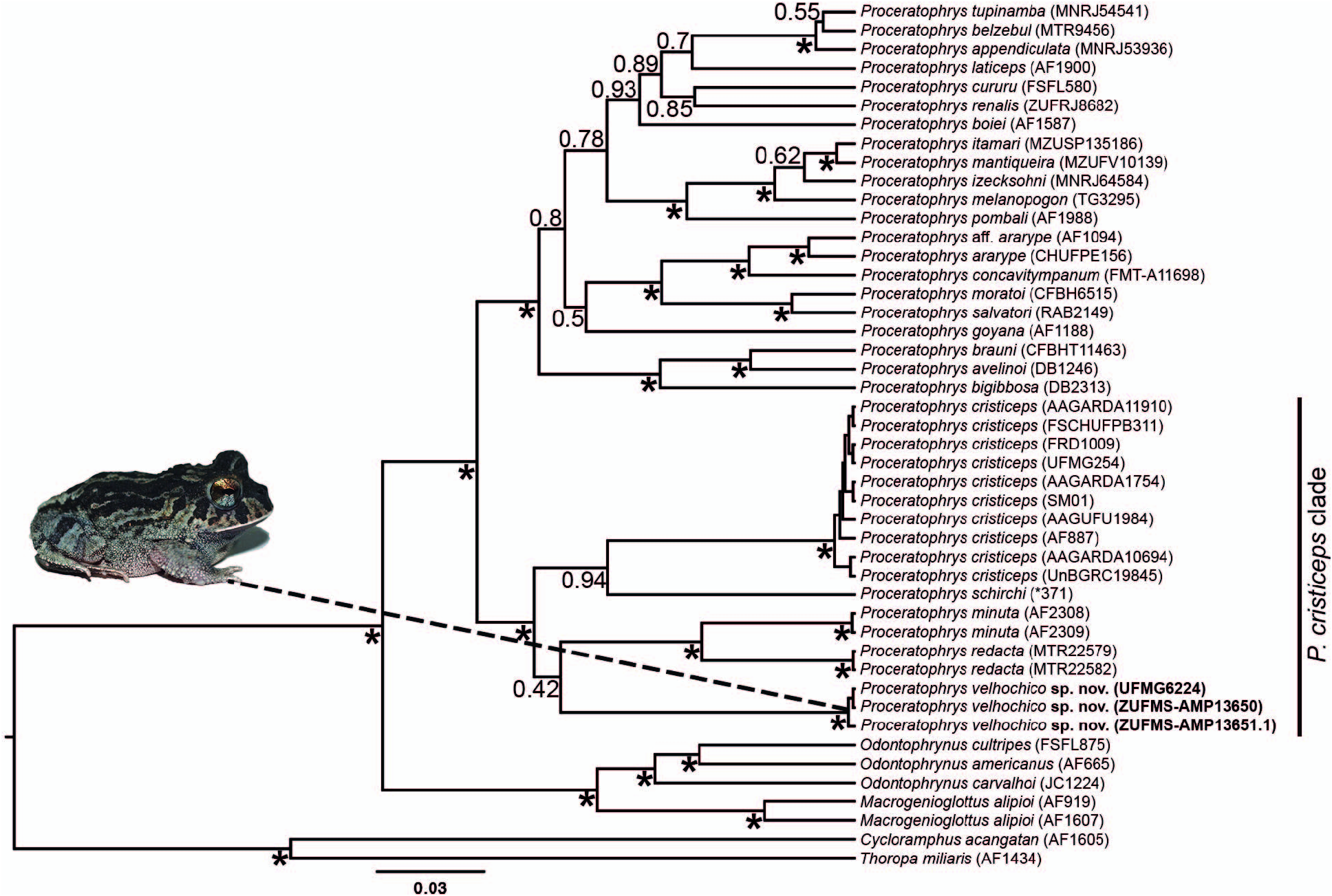
According to the molecular analysis (see below), the new species is more closely related to P. schirchi , P. cristiceps , P. minuta , and P. redacta ( Fig. 5 View FIG ). Besides the differences mentioned above, P. oelhochico sp. nov. also differs from these four species by 1) having a bigger size: SVL 32.1–38.7 mm in males and 37.7– 44.3 mm in females ( P. minuta : 19.2–22.6 mm in males and 25.9– 31.6 mm in females; P. redacta : 26.9–29.7 mm in males and 33.5– 34.9 mm in females; P. schirchi : 30.8 in an adult male; Teixeira et al., 2012), except from P. cristiceps that reaches higher values of SVL (33.1–53.6 in males and 35.1–64.5 mm in females); 2) snout vertical in profile view ( P. cristiceps : obtuse); and 3) by the advertisement call longer (0.327 –0.511 sec) and with more pulses/call (29–46) than P. schirchi (0.210 –0.280 sec, 18–22 pulses/call). With an emission rate of 85.4–96.5 pulses/sec (one recording at 22.6°C), calls of P. oelhochico sp. nov. differ from P. minuta (66.8–73.8 pulses/sec; 19°C) and P. schirchi (76.6– 85.0 pulses/sec; 27°C). The dominant frequency of the new species (1312.5 Hz) is lower than P. redacta (1697–1890 Hz), P. minuta (1980–2070 Hz), and P. schirchi (1640–1810 Hz) and higher than P. cristiceps (861–1125 Hz). Such differences in dominant frequency mirror differences in sizes among species.
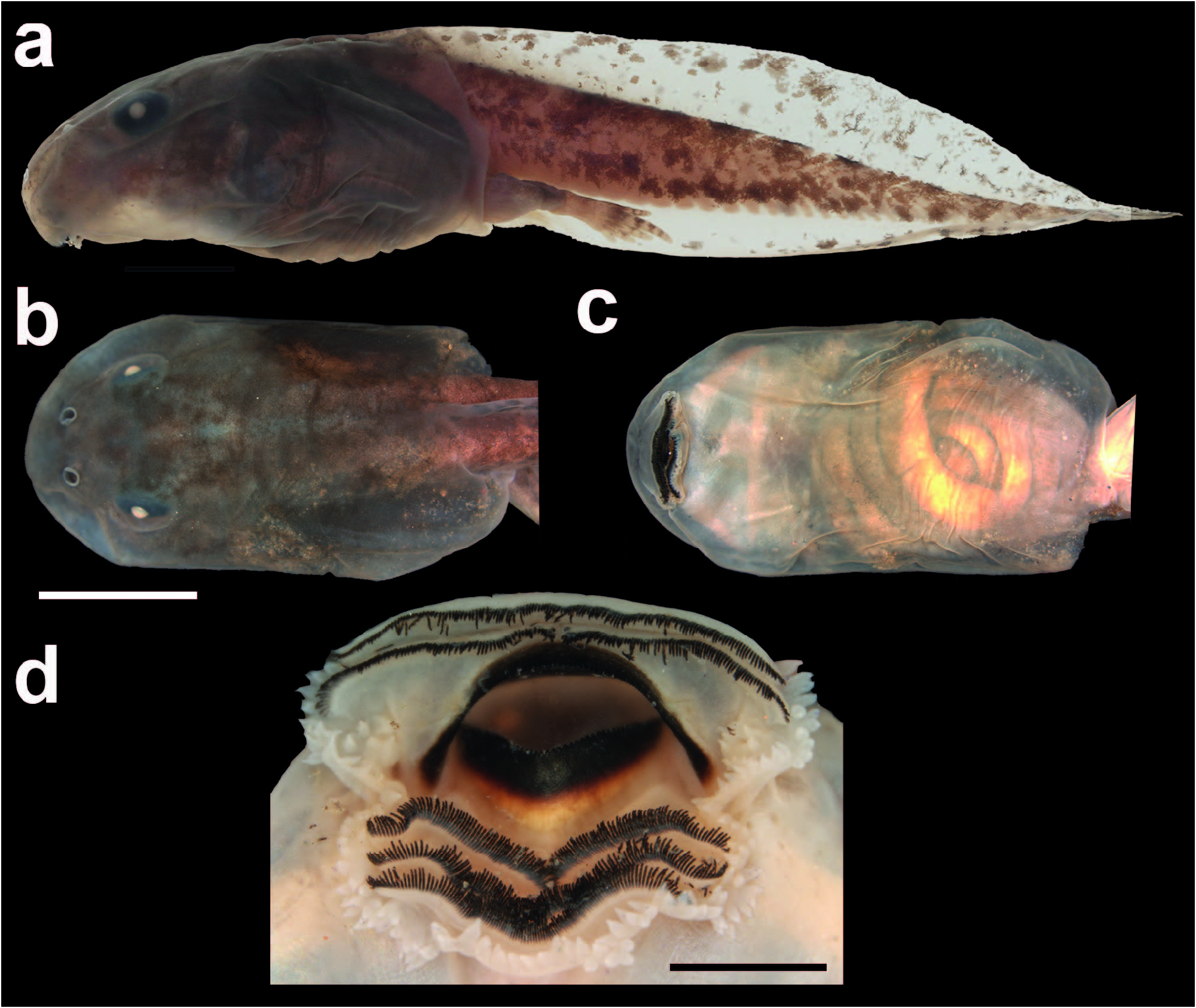
Among the more related species ( P. cristiceps , P. minuta , P. redacta , and P. schirchi ), only the tadpole of P. redacta is unknown. At Gosner stage 39, the tadpole of P. oelhochico sp. nov. reaches a maximum total length of 38.4 mm, distinguishing it from those of P. cristiceps (33 mm) and P. minuta (35.7 mm) at the same development stage. The nares of the tadpole of P. oelhochico sp. nov. are ovoid, bearing a small projection on its inner margin, distinguishing it from the tadpoles of P. cristiceps (projection absent, P. minuta and P. schirchi [nares reniform]). Both marginal and submarginal papillae of the tadpole of P. oelhochico sp. nov. exhibit pointed tips, distinguishing it from P. 128 cristiceps tadpole (rounded), while the tadpoles of P. minuta and P. schirchi lack submarginal papillae. The posterior teeth rows (P1–3) of the tadpole of P. oelhochico sp. nov. are similar in length, distinguishing it from P. minuta (P3 shorter than others). The spiracle inner wall is fused to body but with a small free ridge, distinguishing it from P. minuta (inner wall free from body) and P. cristiceps (inner wall fused to body). The tail pointed tip distinguishes P. oelhochico sp. nov. tadpole from those of P. minuta (rounded to slightly pointed). Moreover, the tail of the P. oelhochico sp. nov. tadpole only exhibits brown and sparse melanophores along its extension, distinguishing it from P. minuta tadpole (tail with 1–5 well-marked broad and black irregular vertical bars). Napoli et al. (2011) and Peixoto et al. (1984) reported that both P. minuta and P. schirchi tadpoles exhibit golden blurs or blotches along the dorsum and flank of the body. Although we did not properly describe tadpole coloration in life, this evident pigmentation was not seen in P. oelhochico sp. nov. tadpoles in life (FMM, pers. obs.). Additionally, the tadpole of P. minuta exhibits reddish faded blurs along its tail, a feature that was also not observed in living tadpoles of P. oelhochico sp. nov.
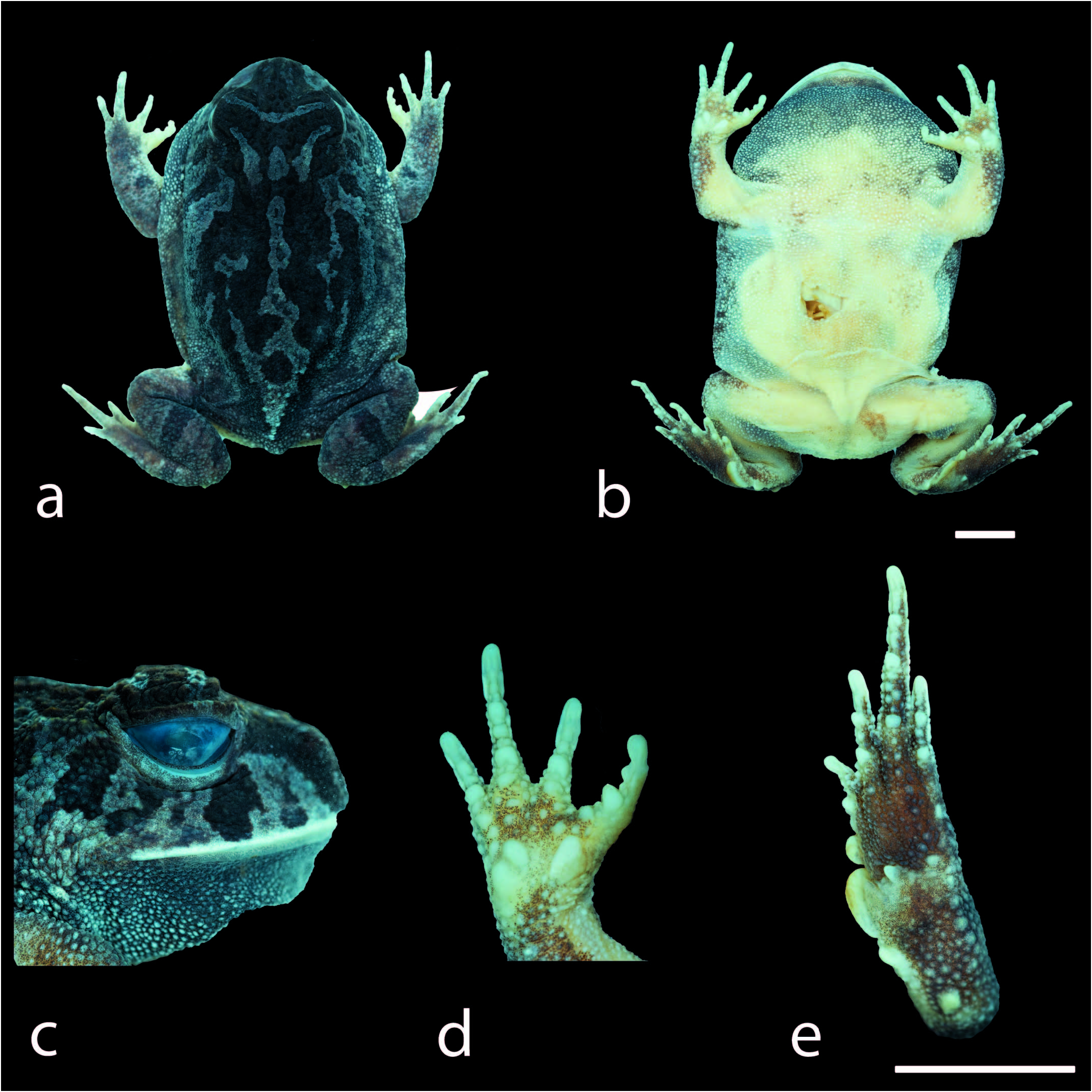
Description of Holotype.—Adult male; aspect robust ( Fig. 2 View FIG ); head wider than long, head length 29% of SVL, snout rounded in dorsal and ventral views ( Fig. 2 View FIG ), and vertical in profile ( Fig. 3a View FIG ); nares elliptical and prominent, canthal crests poorly marked and smooth, no preocular crests; eyes directed anterolateraly, eye diameter 40% of head length and 98% of the palpebral; eyelid without prominent warts; tympanic membrane absent; tympanic annulus absent; tongue cordiform, free posteriorly; vomerine teeth in two short rows between the choannae; vocal sac single, subgular; vocal slits present, lateral to tongue, on the floor of mouth; frontoparietal crests poorly developed; region between frontoparietal crests shallow; absence of inter-ocular and ocular-dorsal ridges of warts. Dorsal surface with few sparse and short warts, one row of rounded and not fused forearm tubercles, reaching the hands; ventral surfaces, except hands and feet, covered by numerous small, rounded, uniform warts. Finger lengths I> II <III> IV ( Fig. 2d View FIG ); interdigital webbing absent; inner metacarpal tubercle large and elliptical; outer metacarpal divided in two oval parts; scarce small rounded supernumerary tubercles; subarticular tubercles large, nearly rounded, grooved anteriorly and posteriorly; nuptial pads absent. Thigh length longer than tibia length, the sum of thigh and tibia lengths 86% of SVL; toe lengths I <II <III <IV> V ( Fig. 2e View FIG ); foot web formula I 2 – 1 II 2 – 1 III 2 – 1 IV 1 – 1 V; inner metatarsal tubercle large, elliptical, spatulated; outer metatarsal tubercle small, rounded, presence of a prominent rounded calcar tubercle; scarce small rounded supernumerary tubercles; subarticular tubercles large, nearly rounded, grooved anteriorly and posteriorly ( Fig. 2e View FIG , 3a View FIG ).
Color in Life of the Holotype.—Dorsal background color grey, maculated with irregular stains dark-grey. Three yellowish bands from eye to upper lip. One transverse light-green band at the inter-ocular region. Two transverse dark-grey bars on the tibia. Fingers, the tip of the toes, and ventral surfaces are cream, with the gular region dark ( Fig. 4 View FIG ).
Color in Preseroatioe of the Holotype.—The dorsal background color changed from light-grey to dark-grey, and the irregular stains became even darker. One transverse dark-grey band at the inter-ocular region. Three light-grey bands from eye to upper lip. Two transverse grey bars on the tibia. Fingers, the tip of the toes, and ventral surfaces are cream, and the gular region is dark ( Fig. 2 View FIG ).
Variation.—Specimens are congruent with respect to morphological characters. The dorsal background color in adult and juvenile specimens varies from grey to cream. The irregular stains on the dorsal surface vary from dark-brown, cream, and reddish (the reddish color remains in preservative; Fig. 4 View FIG ). Adult males have the gular region dark and are smaller than females ( Table 3 View TABLE ).
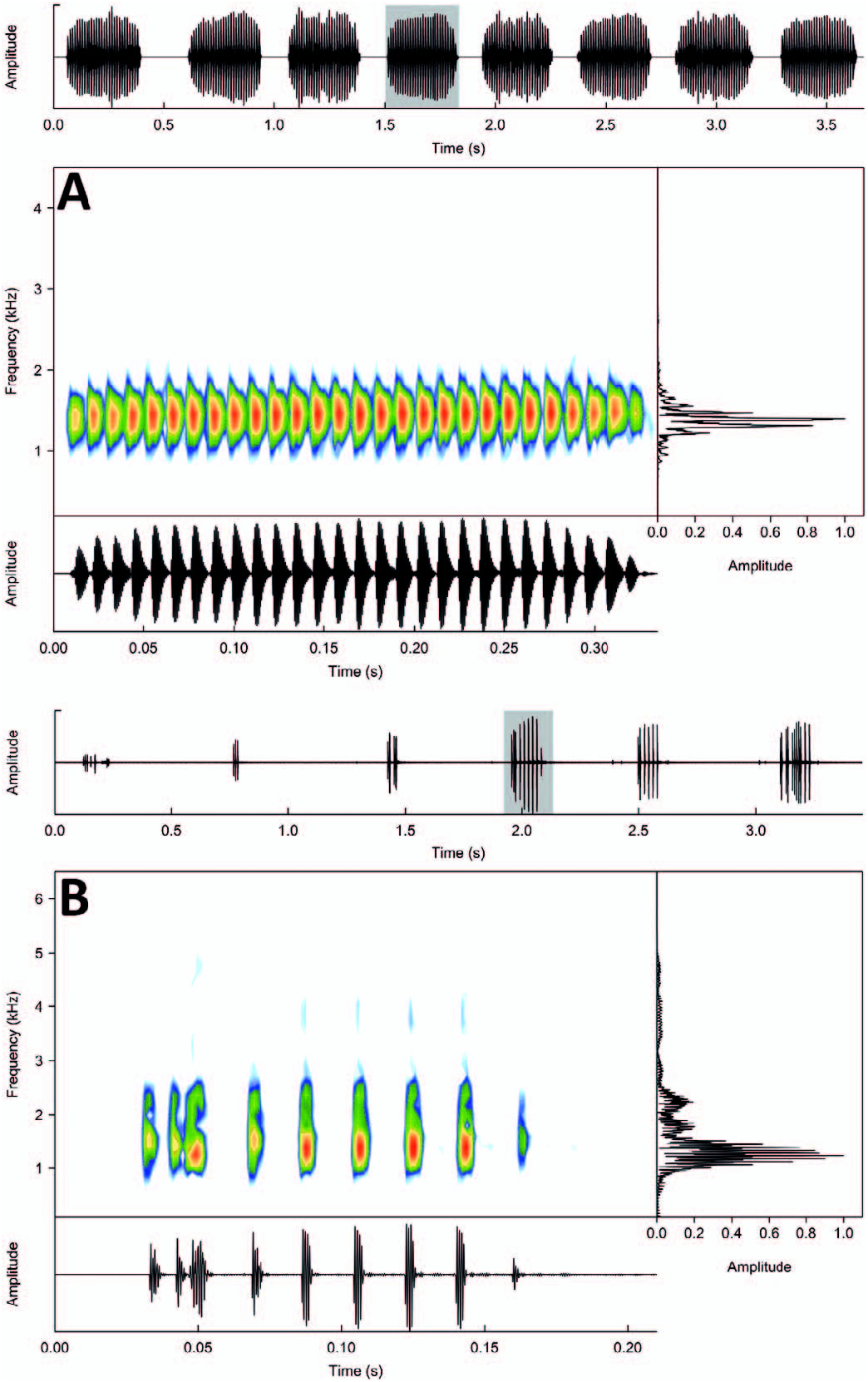
Adoertisement Call.—The advertisement call of Proceratophrys oelhochico sp. nov. is emitted in series of single, multipulsed notes (complete pulses with silent intervals between each one; n = 87 analyzed calls; Fig. 6 View FIG ). The call has a duration of 0.327 –0.511 sec (mean 0.405 ± 0.04) with 29–46 pulses (mean 36.0 ± 3.9), pulse repetition rate from 85.4–96.5 pulses/sec (mean 88.9 ± 2.1), and a dominant frequency of 1312.5 Hz ( Table 4 View TABLE ).
Release Call.—The release call of Proceratophrys oelhochico sp. nov. is composed of a series of single multipulsed notes, emitted randomly (n = 60 calls analyzed; Fig. 6 View FIG ). The call has a duration of 0.014 –0.201 sec (mean 0.083 ± 0.05) with 2–22 pulses (mean 8.15 ± 4.82), interval between pulses 0.003 –0.015 sec (mean 0.010 ± 0.004), pulse repetition rate from 60.2–140.8 pulses/sec (mean 103.7 ± 19.5), and dominant frequency of 1125–1500 Hz (mean 1290.6 ± 109.6; Table 4 View TABLE ).
Molecular Analysis.—The phylogenetic tree recovered Proceratophrys oelhochico sp. nov. nested within a clade composed of P. cristiceps , P. minuta , P. redacta , and P. schirchi with significant support (posterior probability [PP] = 1.0; Fig. 7 View FIG ), although the resolution of internal relationships among them was not significant (hereafter the P. cristiceps clade). Three of these species are distributed within the Caatinga biome and associated phytophysiognomies, and one is endemic to the Atlantic Forest biome ( P. schirchi ), only occurring outside the Caatinga boundaries ( Fig. 8 View FIG ). This clade was recovered as the sister to all remaining species included in the molecular analysis. The phylogenetic relationship of species in this major Proceratophrys clade was mostly recovered with nonsignificant support, except for the previously supported P. biggibosa species group and the P. moratoi clade (including P. moratoi , P. concaoitympanum , and P. ararype ), all recovered with significant support (PP = 1.0; Fig. 7 View FIG ). Additionally, Proceratophrys oelhochico sp. nov. exhibited a minimum of 7% of genetic divergence for the 16S gene and 2% genetic divergence for the rhodopsin gene in comparison to all its congeners ( Table 5 View TABLE ).
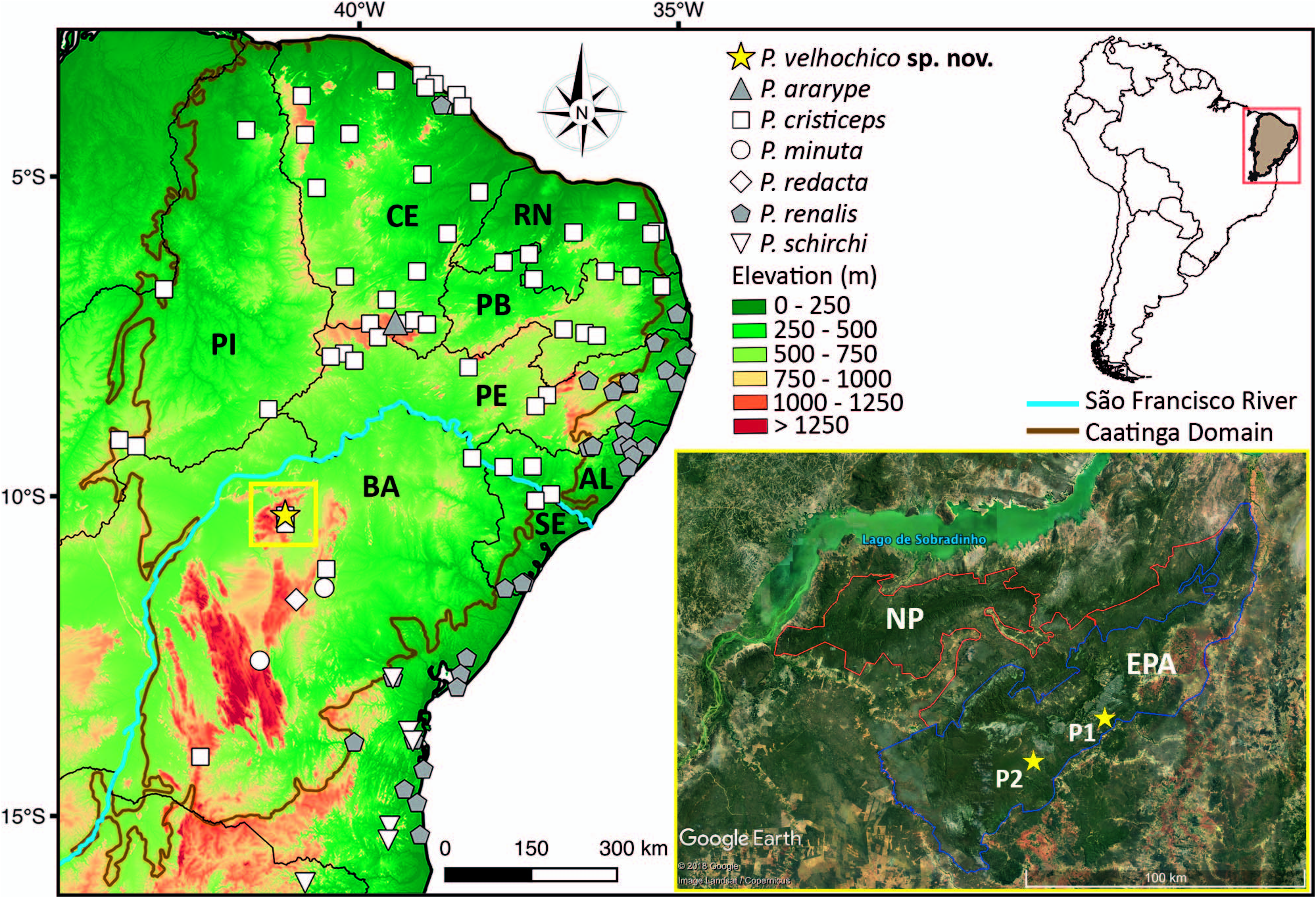
Geographic Distribution.— Proceratophrys oelhochico sp. nov. is only known to occur at two sites (above 800 m a.s.l.) within the APA Boqueirão da Onça , Campo Formoso and Umburanas municipalities, Bahia State, Brazil ( Fig. 1 View FIG ) .
Etymology.—The São Francisco River crosses 2,863 km of the Brazilian territory flowing through Minas Gerais, Bahia, Pernambuco, Alagoas, and Sergipe states. It is one of the most important rivers in Brazil and the largest perennial river in the Caatinga biome, popularly known as Velho Chico (old Chico, which is a popular abbreviation of Francisco). The type locality (Boqueirão da Onça region) is located at the margins of the lower São Francisco River. The specific epithet oelhochico is a noun in apposition.
Tadpole External Morphology.— Proceratophrys oelhochico sp. nov. has an exotrophic, lotic, benthic larvae ( Fig. 8a–c View FIG ; Table 6). At Stage 39, the body is ovoid in dorsal view and slightly dorsoventrally depressed in lateral view (BH/BW = 83%). The snout is rounded in dorsal and lateral views. The eyes are dorsal, laterally oriented. Nares are rounded, dorsally positioned, slightly closer to the eyes than to the snout (END/ESD = 47%), bearing a small and rounded projection on its inner margin. The spiracle opening is sinistral, placed at half of the body’s length and at body’s midline with rounded opening posterodorsally directed; inner wall fused to body with a small free ridge. Vent tube short, median, with dextral opening, right wall displaced dorsally. The tail has about 59% of the total length with pointed tip. The dorsal fin is slightly higher than the ventral one (VFH/DFH = 72%) and the highest portion is at the medial third. The origin of the dorsal fin is at the body–tail junction. The origin of the ventral fin is at the vent tube. The dorsal fin is slightly arched, tapering towards the tail tip; the ventral fin runs in parallel to the tail musculature axis. Myotomes of tail musculature are visible but poorly defined. 130 The oral disc is ventral, laterally emarginated ( Fig. 8d View FIG ). One row of conical marginal papillae around the entire oral disc with a wide gap anteriorly, similar to A1 length. There are 4–5 submarginal papillae on the anterior labium, above mouth commissure. Both marginal and submarginal papillae exhibit pointed tips. Labial tooth row formula (LTRF) 2(2)/3(1); A2 is slightly longer than A1; posterior teeth rows with similar lengths. The jaw sheath is serrated and strongly keratinized, upper jaw sheath arched with long lateral process, lower jaw sheath U-shaped.
In formalin, tadpoles are dark brown with scattered brown blotches along the body dorsal surface. Ventral surface with a lighter brown coloration if compared to dorsal coloration with visible viscera. The iris is black, and pupil is white. Tail musculature and fins with brown and sparse melanophores along its extension, which are denser on the anterodorsal portion. Dorsal fin is more pigmented than the ventral one. Both fins are translucent.
Natural History Notes.—During the field expedition to Campo Formoso municipality (Point 1; Fig. 1 View FIG ), we only collected or heard the new species calling from the ground on sandy banks along the margins of a rocky bottom stream in altitudes above 840 m. We also surveyed for frogs at lower altitudinal areas (between 600 and 700 m) along this same stream (outside the APA Boqueirão da Onça) and only heard or collected Proceratophrys cristiceps in such sites, occurring syntopically with species that are commonly found in the Caatinga, such as Corythomantis greeningi , Odontophrynus caroalhoi , Pleurodema diplolister , Rhinella granulosa , R. jimi , and Scinax x-signatus . It was only after we reached areas above 840 m of altitude (within the APA Boqueirão da Onça) that we first heard the new species in breeding activity, occurring syntopically with P. cristiceps . Males were mostly calling sparsely from each other along the stream (~ 2–3 m away) and stopped calling activity when we approached. Besides P. cristiceps , we also collected Scinax aff. x-signatus calling along the margins of the same stream with the new species ( Fig. 9a–d View FIG ). During this period, it had rained for some days before we surveyed the region.
At Umburanas municipality on 4 November 2020 (Point 2; Fig. 1 View FIG ), we collected frogs a day after a torrential rain in a rupestrian field area above 985 m of altitude (also within the APA; Fig. 9e–f View FIG ). We observed around 10 individuals calling in a flooded, open anthropized area (~ 5–8 m away from each other) amidst grassy vegetation ( Fig. 4d View FIG ). At this site the new species was found occurring syntopically with Leptodactylus oastus , L. troglodytes , Phyllomedusa bahiana , Scinax pachycrus , and Rhinella jimi . Near to the reproductive site we found an individual in a disabled road, and it displayed a stiff-legged behavior when we handled it ( Fig. 4f View FIG ). At 2100 h we left the site, and some individuals were still in calling activity. On 8 November 2020, we returned to this same site and found one female (ZUFMS-AMP 13949) along the route to the reproductive site. We stayed at the location from 1900 h to 2100 h and did not hear any P. oelhochico sp. nov. males, only the other species mentioned above. All the females collected on 4 and 8 November 2020 were ovigerous. At this same site, on 8 February 2021, six males 132 started to call at 1950 h (~ 5–10 m away from each other), syntopically with Scinax pachycrus and Corythomantis greeningi .
In accordance to the above mentioned facts, we conducted four other field trips, including both dry and rainy seasons, in lowland areas in the region of Umburanas and did not record Proceratophrys oelhochico sp. nov., only populations of P. cristiceps . Such evidence suggests that the new species could be restricted to high elevation areas of Boqueirão da Onça and that it is likely an explosive breeder and uses lentic and lotic habitats flexibly. However, more field efforts will be needed to ascertain the distribution, conservation status, and natural history observations of P. oelhochico sp. nov.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.