
Carinacauda stormi, Leonard, William P., Chichester, Lyle, Richart, Casey H. & Young, Tiffany A., 2011
|
publication ID |
https://doi.org/ 10.5281/zenodo.276654 |
|
DOI |
https://doi.org/10.5281/zenodo.5685274 |
|
persistent identifier |
https://treatment.plazi.org/id/039187F6-FFBD-FFAB-FF36-66A69BA6FC3B |
|
treatment provided by |
Plazi |
|
scientific name |
Carinacauda stormi |
| status |
sp. nov. |
Carinacauda stormi View in CoL new species
Fig. 5 View FIGURE 5 , 6 View FIGURE 6 , 7 View FIGURE 7
Types. The holotype ( CM 104044 View Materials ), taken on 15 November 2008 from the Upper Blue River Watershed, and one paratype ( CM 104023 View Materials ), taken on 0 5 November 2009 from the Upper Canyon Creek, were collected by T. Young from Douglas-fir needle duff litter ( Pseudotsuga menziezii ) on the Willamette National Forest, Linn County, Oregon. An additional paratype ( CM 104045 View Materials ) was collected on 15 April 2010 by T. Young in the Canyon Creek Watershed, Willamette National Forest, Linn County, Oregon.
Etymology. The species name honors Dr. Robert M. Storm, professor emeritus of natural history and vertebrate zoology at Oregon State University.
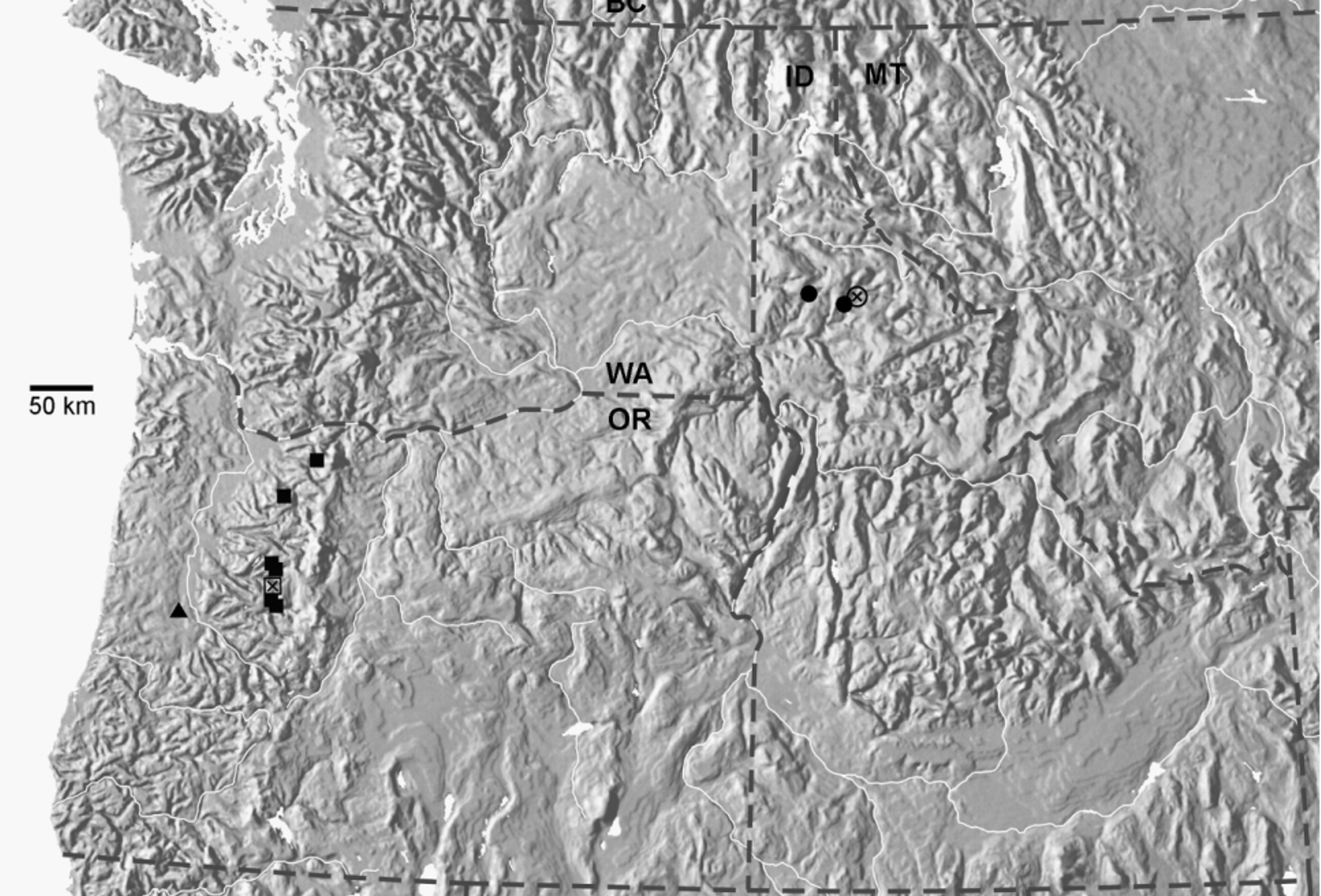
Distribution. This species was detected at more than 50 localities at elevations ranging between 610 to 1190 m asl in Willamette and Mt. Hood National Forests in Clackamas, Marion, Linn and Lane counties, Oregon ( Fig. 1 View FIGURE 1 ).
Description. Size: Very small, extended length of adults in movement 12 to 15 mm; length reduced by approximately 40% after preservation.
External features: As in Securicauda except the following: mantle 58 to 64% (mean=61%; SD=0.04; n=15) of the extended body in movement and 55 to 80% (mean=77%; SD=0.07; n=12) of preserved specimens; pneumostome located in the middle (sometimes slightly posterior) of right side and bottom of mantle; tail 20 to 27% (mean=24%; SD=0.03; n=15) of the extended body in movement.
Coloration: Head pale-gray or translucent tan-white; tentacles dark-brown/gray. Neck semi-translucent; adults with a distinct white patch on right side. Mantle tan or pale-gray with light-blue flecking and dark-brown or gray irregular blotches, with a relatively broad lateral stripe on each side. Pneumostome located below lateral stripe and surrounded by light-gray “halo.” Sides below mantle pale-gray, with or without dark-brown or gray markings. Tail pale- to dark-gray with fine white flecking and dark-brown or gray markings coincident with furrows; usually with a pale-gray stripe on mid-dorsal keel. Sole pale-gray with scattered white flecks. Mucus clear.
Internal features. Shell: Similar to S. hermani in both position and composition.
Jaw and Radula: Similar to S. hermani in structure.
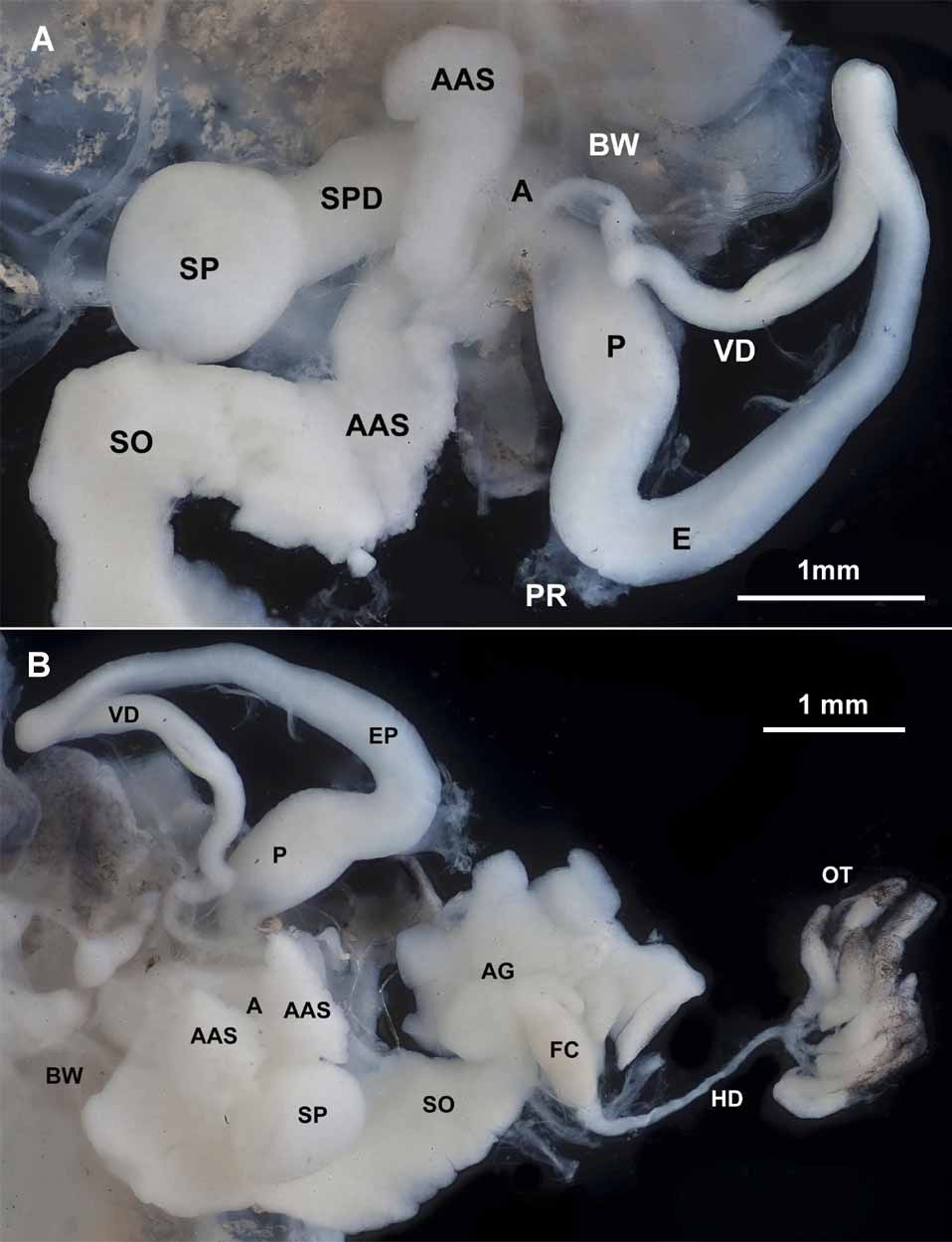
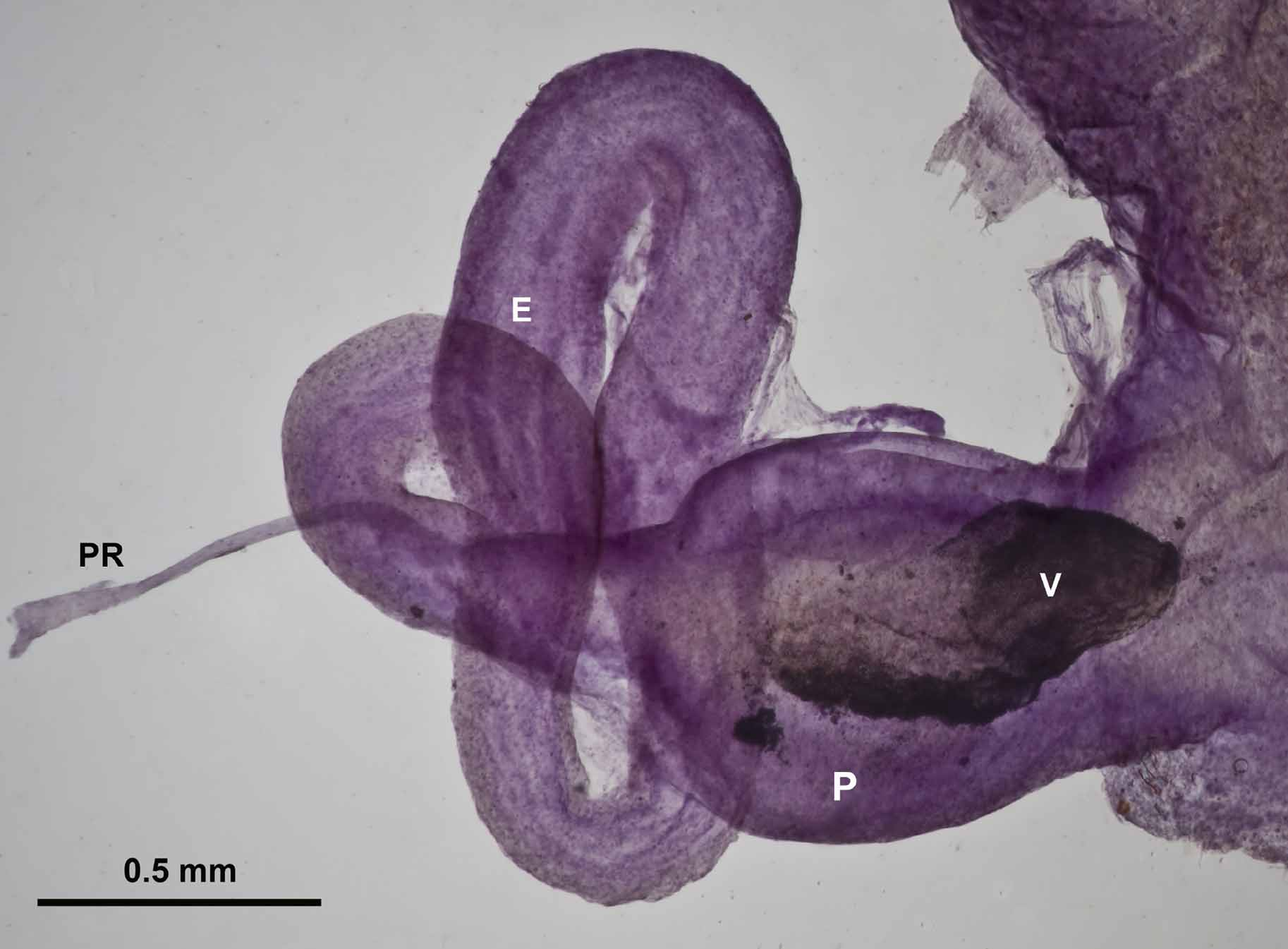
Reproductive System: Ovotestis consisting of 12 or more lobules; located on midline just under posterior margin of diaphragm and partially surrounded by the digestive gland. Hermaphroditic duct of moderate length and thickness; leading to fertilization chamber, which is largely surrounded by the large albumen gland. The whitish, loosely coiled spermoviduct leads to the tubular free oviduct that opens into the atrium. The spermathecal duct, which arises near junction of free oviduct and atrium, generally becomes thick in sexually mature individuals; in two of our individuals the duct remained slender for at least a portion of its length. Spermatheca large and globular. The slender vas deferens arising from spermoviduct; traveling across the free oviduct and connecting with the long, thick and highly coiled epiphallus. Penis retractor muscle inserting in wall of epiphallus just before duct joins penis apex. Epiphallus internally connecting with a large verge that is about half as long as the penis. In sexually mature individuals the distal portion of the verge is so opaquely white that it can be seen through the translucent wall of the penis (in Fig. 7 View FIGURE 7 these opaquely white areas show up as densely black). Penis opening into atrium, which also receives the free oviduct and spermathecal duct. The atrium also bears a pair of accessory structures of unknown function. In immature animals, these structures resemble the cotyledons on a bean sprout. As animals mature the pair becomes much larger and more complex. They are sufficiently opaquely white for the more lateral member of the pair to be seen through the thin body wall. Examination under a compound microscope failed to reveal any fine detail—no darts, or ducts or muscle bands that might suggest a special function.
Buccal and tentacular retractor muscles: Similar to the architecture of S. hermani .
Digestive system: As in S. hermani the crop and the pair of digestive glands are the major feature of the digestive system. The intestine is short and oriented as in S. hermani .
Natural history. This species is currently known from montane areas of the Oregon western slope Cascades where Douglas-fir ( Pseudotsuga menziesii ) is a dominant overstory species and minor tree species include Western Hemlock ( Tsuga heterophylla ) and Vine Maple ( Acer circinatum ). We located approximately 100 individuals at more than 50 sites in Willamette National Forest; all individuals were found in Douglas-fir–Western Hemlock forests where needle litter was almost exclusively Douglas-fir at the microsite. Forest age class did not seem to be a factor in detecting this species; detections occurred in forests 25 years to over 150 years in age. Areas where down wood retained pockets of moisture and where Vine Maple leaves formed a layer to hold moisture are preferred habitats. In addition, small topographic depressions where water collects or saturation occurs as the rainy season develops appear to be important microhabitat features. Detections occurred almost exclusively in semi- to fully saturated needle litter between the most recent and previous year’s needle-duff layers (approximately 2.5 to 5 cm below the surface). Shrub, fern and moss layers were lacking in the areas where detections occurred. This species was rarely detected in forest stands where either Incense-cedar ( Calocedrus decurrens ) or Western Redcedar ( Thuja plicata ) was a dominant tree species. Though it has been found at the margins of small streams, this species is not associated with seeps or wetlands.
Detections have occurred primarily in the fall, but recent spring surveys have detected adults and juveniles in conditions where litter was still saturated due to snowmelt. Four specimens have been successfully kept in captivity for 24 months (November 2008 to November 2010) and over 20 collected in November 2009 are currently in captivity to document biological and reproductive behavior. Eggs were documented in February 2009 and March 2010; however, none of the eggs produced hatchlings. The clutch in 2010 contained 12 eggs that were semi-translucent white, oblong and approximately 2 mm long.
Comparative anatomy. We place Carinacauda stormi in the Arionidae on the basis of its ribbed jaw, which is the only character it shares with all other arionids. It differs from Securicauda hermani by possessing an atrium and paired accessory structures and a well developed, coiled epiphallus. It differs from Kootenaia and Prophysaon by virture of its keeled tail and from all other genera in Table 1 View TABLE 1 by its divergent buccal and tentacular retractors.
Additional comments. We suggest the common name Cascade Axetail in recognition of the Cascade Mountains where the species resides and to reflect its hypothesized close relationship with Securicauda .

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |