
Sepedonella nana
|
publication ID |
https://doi.org/10.5281/zenodo.206551 |
|
DOI |
https://doi.org/10.5281/zenodo.6193176 |
|
persistent identifier |
https://treatment.plazi.org/id/0391B42E-FFBF-FF92-78FA-FF2BFC0AF9F3 |
|
treatment provided by |
Plazi |
|
scientific name |
Sepedonella nana |
| status |
|
Sepedonella nana View in CoL is a small fly, 3.5–6 mm long. It is one of the most abundant sciomyzid species collected in Benin. The species has been described by Verbeke (1950) from specimens collected in different National parks of the Republic of the Congo. The male type and paratypes (both sexes) are in the Institut royal des Sciences naturelles de Belgique, Belgium, Brussels. Later, Verbeke (1961) mentioned its presence in Lesotho; Knutson (1980) listed Chad, Ethiopia and Nigeria; in addition, besides our collection in Benin we caught S. nana View in CoL in Togo and Nigeria.
Adults were captured in all our temporary and permanent prospected localities and in additional freshwater areas that we visited irregularly. Among all sciomyzids collected ( Table 1), the species represented 20.35% of the captures. It was associated with the abundant Sepedon ruficeps View in CoL , S. trichrooscelis and S. ornatifrons Adams 1910 View in CoL . Sometimes it was collected with the uncommon Sepedon lippensi Verbeke 1950 , S. umbrosa Verbeke 1950 View in CoL , and exceptionally with the rare species S. ( Mesosepedon) knutsoni View in CoL .
Species q captured individuals ( 1998–2001) Relative abundance q/Q% Sepedon ruficeps View in CoL 2 877 38.80 Sepedon trichrooscelis 2 284 30.80 Sepedonella nana View in CoL 1 509 20.35 Sepedon ornatifrons View in CoL 690 9.31
Sepedon lippensi 27 0.36
Sepedon umbrosa View in CoL 24 0.32
Sepedon knutsoni View in CoL 4 0.0 5
Total captured Q 7 415 100%
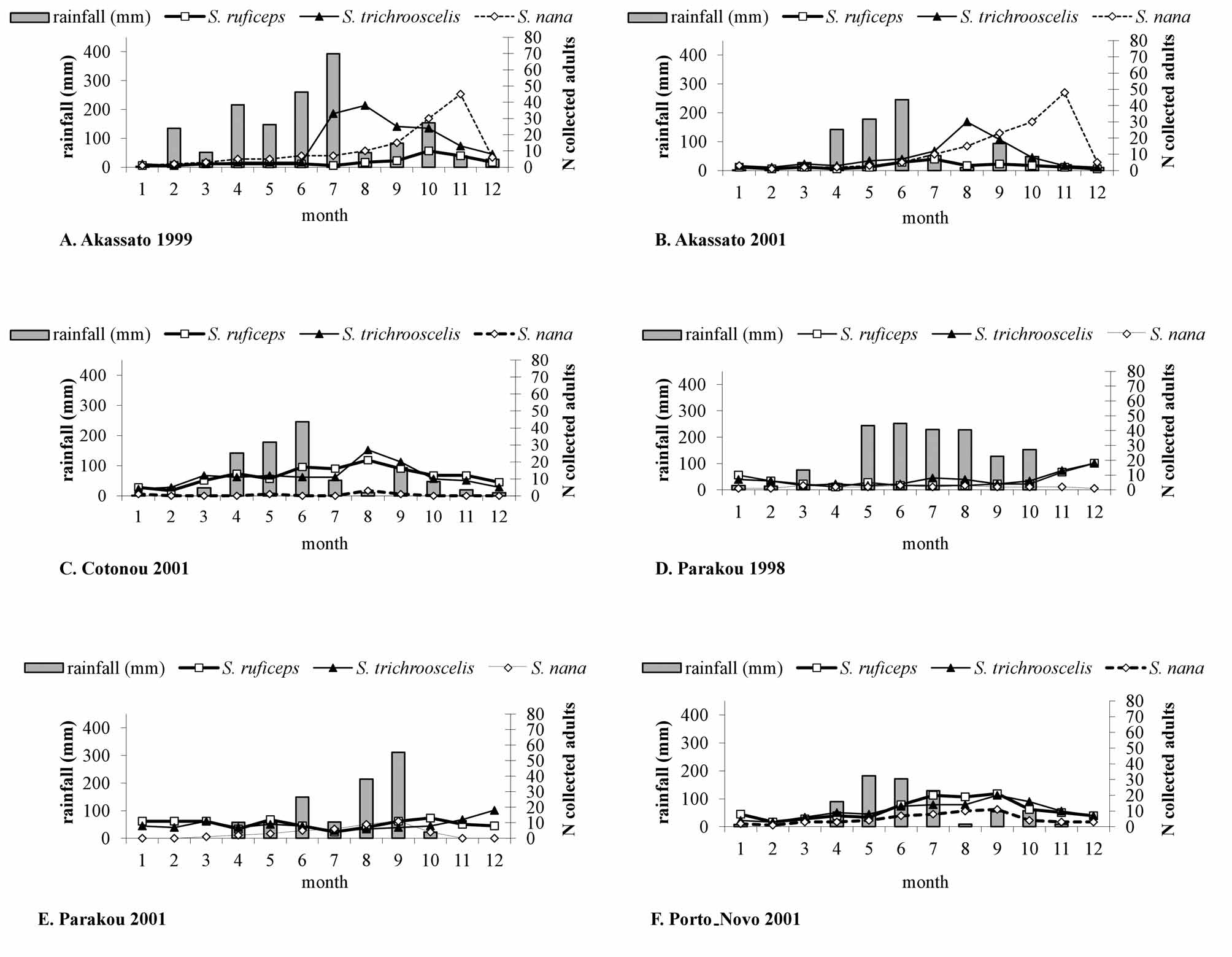
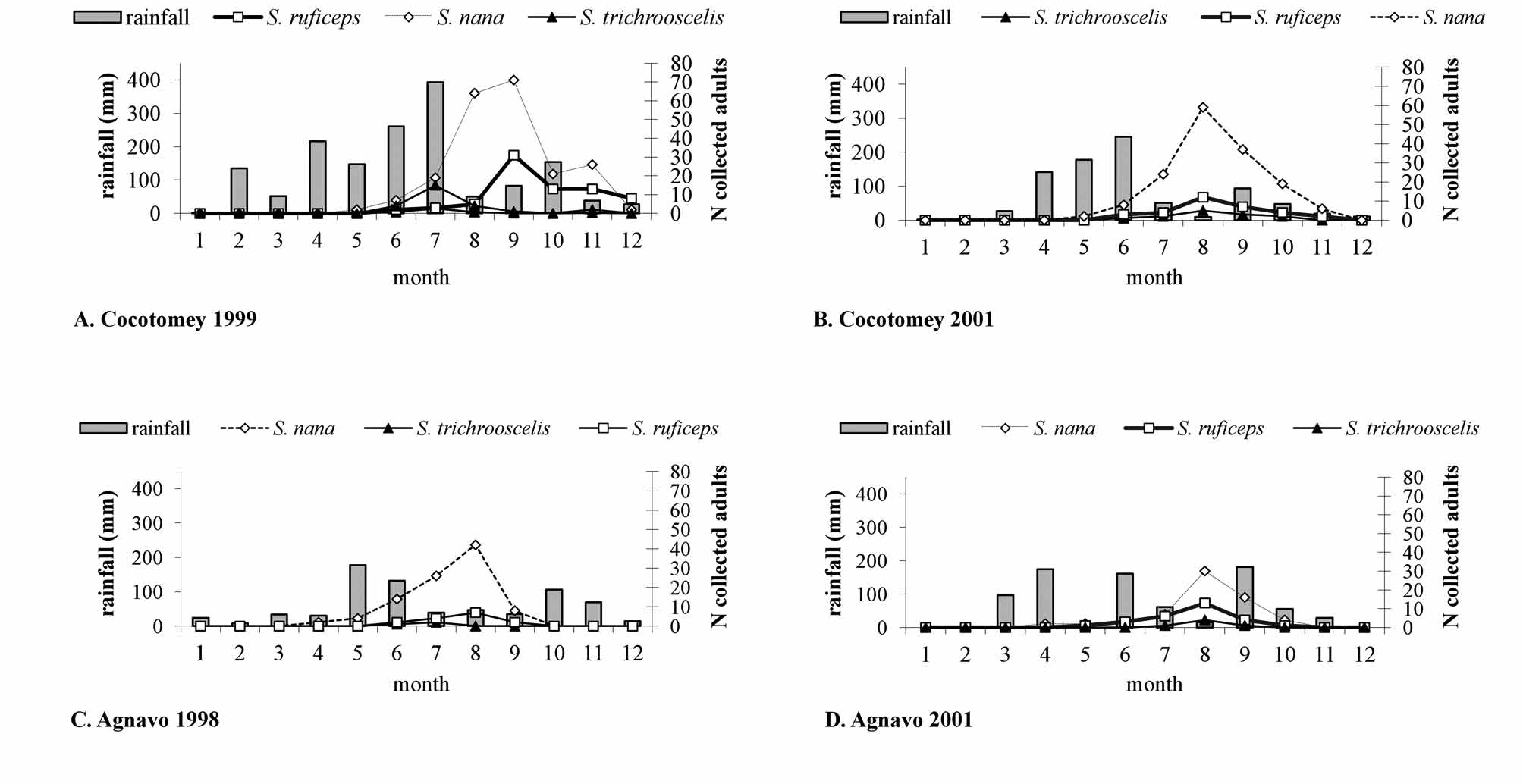
Population dynamics ( Fig. 2 View FIGURE 2 , 3 View FIGURE 3 ). Each year, Sepedonella nana View in CoL was collected with species cited above. But, as Sepedon ruficeps View in CoL and S. trichrooscelis were constantly present or the dominant species, we added them on our population dynamic graphs.
In Akassato, the number of S. nana View in CoL adults collected was low from January to June (2–7 individuals), rising steadily in subsequent months to reach a peak in October-November (48–50 individuals) before decreasing but never was zero ( Fig. 2 View FIGURE 2 A, B). In relation to rainfalls, the peak was subsequent to the period of heavy rain. In Cotonou, S. nana View in CoL adults were present sporadically during the year but always in very low numbers, probably because of the impact of wastewaters from the city that partially irrigate the biotope ( Fig. 2 View FIGURE 2 C).
In Parakou, as the site was always inundated, the submerged surface varied following the seasons. S. nana View in CoL adults were captured at low densities when the level of rain was high as during the year 1998 ( Fig. 2 View FIGURE 2 D). However, in 2001, the rainy regime varied over the year; a minor peak of flies followed the high rains located in August–September ( Fig. 2 View FIGURE 2 E). In Porto Novo, the evolution of the S. nana View in CoL adult was closed to the results of Parakou; but the number of collected adults was higher throughout the year and the rainy season more spread out and mainly localized early in the year ( Fig. 2 View FIGURE 2 F). All these weather variations illustrate the correlation between the fly population dynamics and the rainfalls.
In temporary aquatic biotopes, Agnavo and Cocotomey, the adults captured were always lower and the annual profile of the S. nana View in CoL population was constantly the same. The main feature was the presence of a unique peak after the rainy period ( Fig. 3 View FIGURE 3 ). Only the annual position of the peak was advanced or delayed depending on the variability of the rainy periods.
Discovery of the natural prey of the larvae. Because most reared sciomyzid larvae are malacophagous, we temped to rear S. nana View in CoL larvae on the autochthonous mollusc species, brought to the laboratory. Of more than 200 first-instar larvae tested, only two barely tried to consume on a crushed Physidae View in CoL . They died before reaching the second instar. All of the following laboratory attempts were unsuccessful. No larvae were seen attacking Physidae View in CoL or any other mollusc species, alive or crushed, young or old, aquatic or terrestrial. The eggs of these molluscs were also ignored by the larvae. Fortuitously, in the laboratory we discovered a few S. nana View in CoL puparia floating in some aquaria which contained only water carried from the biotopes, i.e. small vegetal fragments on the surface, and a thin layer of mud on the bottom. These aquaria did not contain any snails.
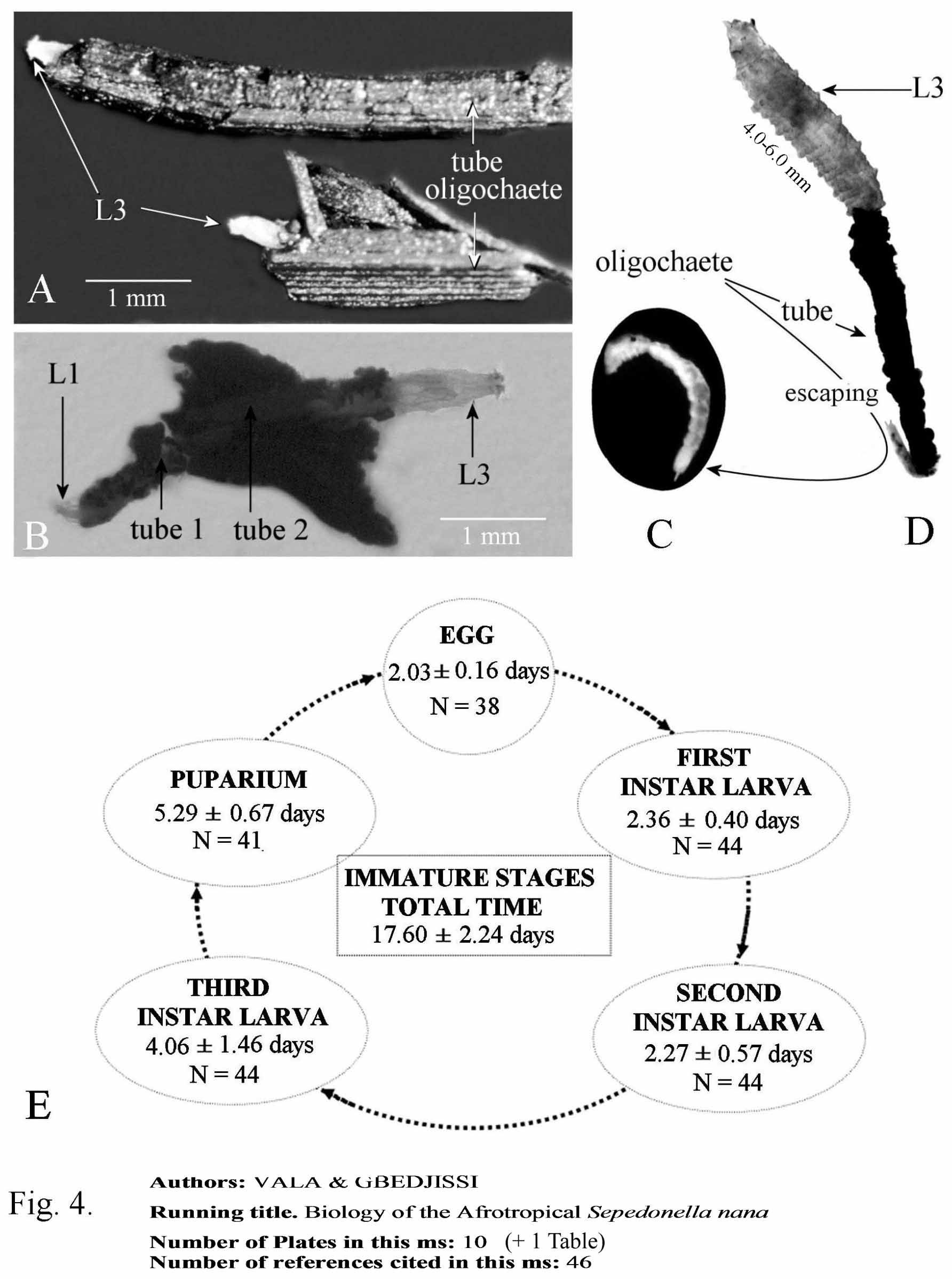
We examined closely the aquaria that contained these puparia, and we found one living second-instar larva, lying on a fragment of a floating plant. However, on close inspection it was found that some small plant fragments were moving on the surface! Upon closer dissecting scope inspection we found that plant fragments were associated with the presence of the small annelid Aulophorus furcatus (Müller) , Oligochaete Naïdidae . In fact, this annelid, 3–7 mm in length, builds a protective tube open at both ends which is composed of leave, stem, and root fragments of plants ( Fig. 4 View FIGURE 4 A–D). Its posterior end shows two furca. It moves with the tube and retracts inside when attacked. Since this oligochaete was observed to be associated with the larval stage of S. nana View in CoL we felt that it may be the potential prey of the S. nana View in CoL larvae. To check this hypothesis, neonate S. nana View in CoL larvae were transferred into Petri dishes along with 10 living A. furcatus , and observed twice each day. The results confirmed that the worms were indeed predated upon by the S. nana View in CoL larvae. In total over 500 rearings confirmed the life-cycle of S. nana View in CoL as using this annelid exclusively as prey. When offered a choice of various mollusc species and this oligochaete, only the worms were attacked and consumed, and would and die in the presence of snails when offered as the sole food source. Observations in the field and on samples brought back to the laboratory confirmed that larvae were found predating on this oligochaete and living in the protective tubes.
Biology. Reproductive behaviour and laying eggs in laboratory conditions. The abdomen of both sexes in ventral view has three distinct forms: very inflated, slightly convex and slightly concave ("hollow" form). In the laboratory, only females with inflated and convex abdomens lay eggs. These two abdominal forms widely predominate among individuals captured during the raining season. The slightly concave abdomen occurs more frequently during the dry season and accounts for up to 30% of collected flies, which may represent sexually immature individuals, those in quiescence (?), or females at the end of the laying period. In nature, as well as in the laboratory, females lay eggs on emerged leaves in lines of 2 to 50 eggs stuck to each other on their sides. From our laboratory data on 30 females, each lays about 125– 250 eggs.
Larval movement and feeding. Sepedonella nana larvae are carried passively by water movements or swim on the surface by their own means. In many instances, larvae were found in nature swimming beneath the water surface with their posterior spiracular disk in direct contact with the air. Larvae moved either by crawling on the water surface or swimming against the slow currents of the water. In the latter case, the larvae contracts the anterior end, then swiftly extended themselves, and then alternately swung their posterior end from right to left and vice-versa. These movements often result in encounters with vegetal fragments and among them, oligochaete prey hidden inside their sheltering vegetal tubes. We did not enumerate the prey density, though it seems high in comparison with the number of snail-predating living sciomyzid larva and fly adults collected in all stations.
Once in contact with a prey’s tube, the larva attacked quickly stretching over the tube and searching for the anterior orifice to attack the oligochaete worm inside. When the annelid was touched by the larva, it rapidly retracted into the tube; the larva followed it, and progressively crawled into the tube while consuming the worm. The posterior disk of the larva was always extruded outside the tube ( Fig. 4 View FIGURE 4 A). Ingested food was visible through the transparent digestive tube of the larva, which did not stay in the prey’s protective tube for more than one hour. Sometimes, the same oligochaete was attacked by two larvae, one from each end of the tube. In many instances, several A. furcatus were grouped on a large vegetative fragment, thus allowing us to observe 1– 3 larvae alongside of each other ( Fig. 4 View FIGURE 4 B).
Larval attacks may be unsuccessful. Aulophorus furcatus avoided assaults by sciomyzid larvae by exiting quickly out the posterior end of the tube which remained floating on the water surface ( Fig. 4 View FIGURE 4 C, D). The exposed oligochaete then fell to the bottom of the Petri dish and the larva then desisted. The worm would then quickly gather plant fragments around its body and form its protective tube. Oligochaeta and S. nana larvae predominantly live at the water surface where the larval predation of the worm is carried out. The larvae of S. nana were observed to not attack or feed under the water. However, the larvae may continue to feed under water for about 5–10 minutes if they are dragged to the bottom of the Petri dishes with the worm.
A single larva of S. nana fed on 10–21 (mean 14.52, n = 132) oligochaetes during its life. The tube diameter is also a determinant factor; it must be sufficiently wide to enable entrance of the S. nana larva, which may be trapped inside if it is too narrow ( Fig. 4 View FIGURE 4 D). Unsuccessful attempts may also increase the number of oligochaetes attacked, partially consumed, or killed as worm fragments were seen to be strewn on the bottom of rearing Petri dishes. Here, we observed asexual regeneration of many Aulophorus furcatus fragments into complete worms as noted by Foulkes (1953), due to the mobilization of neoblasts present in each free fragment. Incidentally or intentionally, this phenomenon can easily be used to maintain a permanent reserve of worms in the laboratory.
Duration of immature stages ( Fig. 4 View FIGURE 4 E). As all sciomyzid species, the immature stages regroup the egg, first-, second- and third instars, and the puparium. The total length of the cycle varied from 14 to 21 days. The embryonic period was very short with hatching occurring two days after being laid. First instar larvae moulted into second instars after 2 or 3 days. The second instar lasts 2–3 days, the third instar 4–6 days and the puparial period 5–6 days. Thus, the cycle is very rapid and reflects in accordance with the multivoltine character of S. nana , which adults are present throughout the year, mainly in permanent ponds, marshes or similar permanent aquatic biotopes.
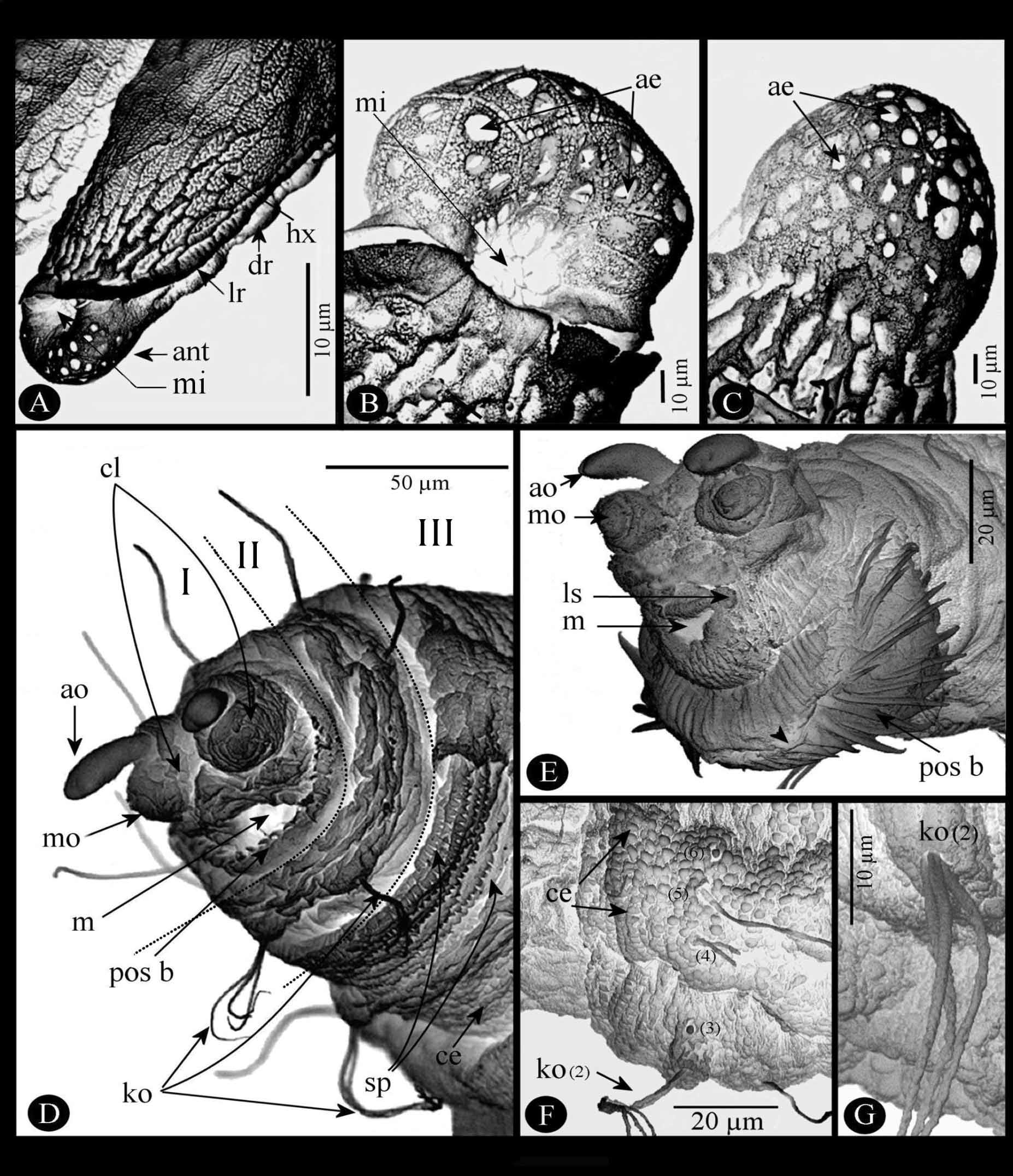
Description of immature stages. Egg ( Fig. 5 View FIGURE 5 A–C). Length 0.80–1.05 mm; greatest width 0.20–0.23 mm. Whitish just when hatched, light brown afterward. Chorion with a reticulate surface of joined hexagonal areas (hx) with 10–15 rounded asperities in each ( Fig.5 View FIGURE 5 A). Two sub-dorsal longitudinal ridges (dr) and two sub-lateral longitudinal ridges (lr) delimiting four distinct areas ( Fig. 5 View FIGURE 5 A): one dorsal, slightly concave, strongly reticulate; one very narrow lateral area on each side between the sub-dorsal and sub-lateral ridges; one large convex ventral area partially covered by a glue that fixed the egg onto vegetation or other support. Anterior end (ant), more or less rounded ( Fig. 5 View FIGURE 5 A, B), greatest diameter 75.00–82.50 μm, length 62.5 μm, micropyle (mi) ventral, about fifty aeropyles (ae). Posterior end ( Fig. 5 View FIGURE 5 C), subspherical, greatest diameter 80.00–91.66 μm, length 86.66–89.00 μm well delimited by a circular-like ring, presence of about one hundred aeropyles (ae). (Based on 5 specimens).
Larvae. The three instars are very similar in general shape, morphological structures, and sensorial features. Twelve segments (I–XII) compose the body: 1 cephalic, 3 thoracic (th1-th3) and 8 abdominal (ab1-ab8). The main differences are in body length, coloration, size, and form of the cephalopharyngeal skeleton, and some characters absent in first-instar and present in second- and third-instars as the anterior spiracles or the structure and size of the posterior spiracular disc.
1. General description. Subcylindrical, anterior end more acute. Whitish at hatching becoming greyish after. Integument transparent. Body surface extensively covered with many round cuticular expansions ( Fig. 5 View FIGURE 5 D, F, ce), regularly distributed.
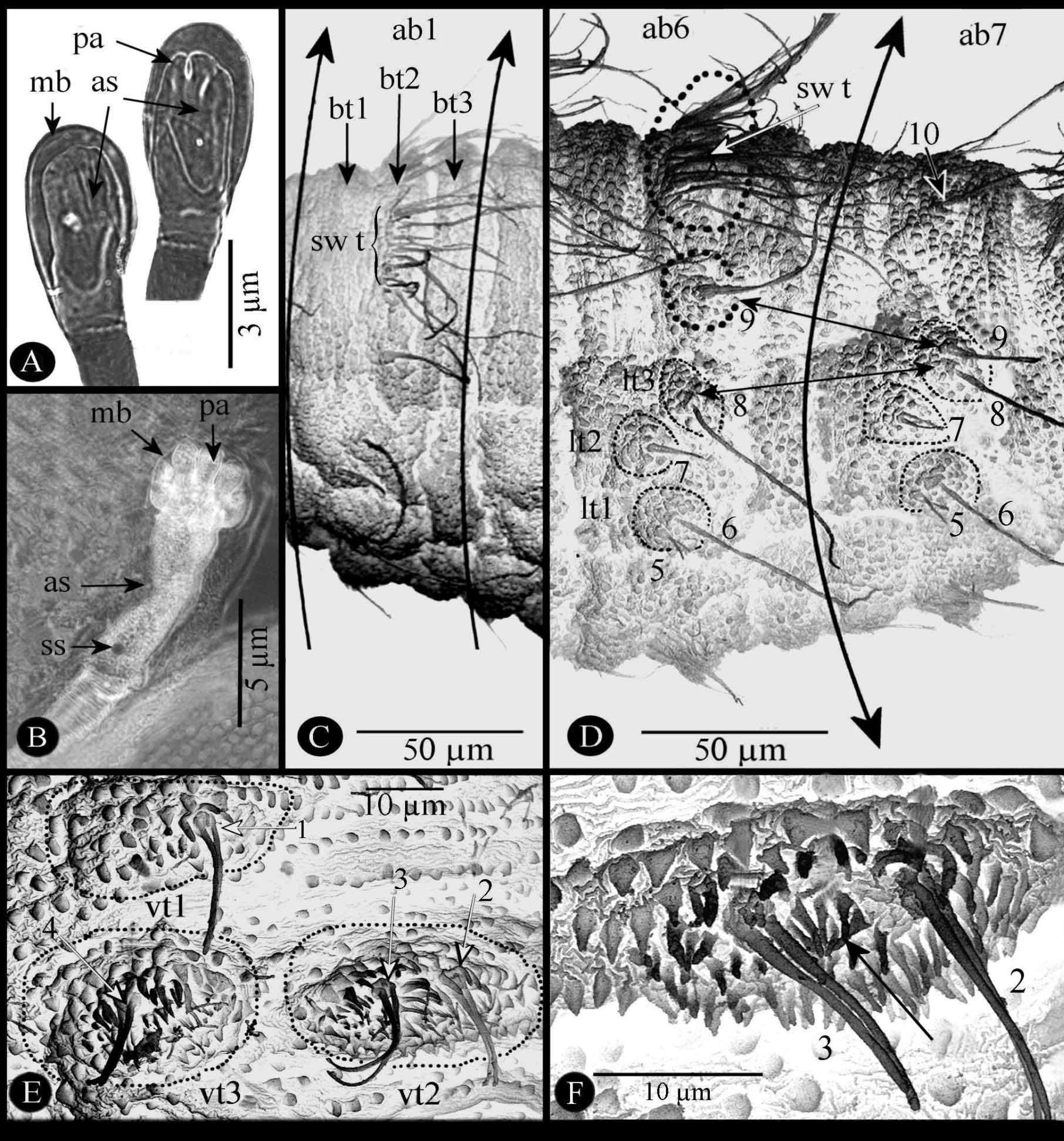
Segment I (cephalic segment) ( Fig. 5 View FIGURE 5 D, E) apically bilobed (cl), each with one prominent antennal organ (ao) anteriorly directed and one elliptical maxillary organ (mo) with some sensilla. Mouth (m) ventrally positioned with two labial sensilla (ls) on each side ( Fig. 5 View FIGURE 5 E). Posteroventral border with a large postoral spinulete band (pos b) ( Fig. 5 View FIGURE 5 D, E), these spinules very long ( 20–31.43 mm length) with apex pointed, not divided.
Segments II–IV (thoracic segments) easily identified by presence of one ventral pair of Keilin organs (ko) ( Fig. 5 View FIGURE 5 D, F, G) composed of three long chaeticum-type sensilla erected from unique annular hole. Anterior margin of segments III and IV (th2, th3) with a transversal spinulated ventral band (sp) of 5–6 rows of short spinules (3.41 µm long) ( Fig. 5 View FIGURE 5 D), spinule apex often bifid. One anterior spiracle (as) on each lateral segment II only on second- and third-instar larvae ( Fig. 6 View FIGURE 6 A, B).
Segments V–XI (abdominal segments ab1-ab7), each with three transverse bands on dorsal surface (bt1-bt3) ( Fig. 6 View FIGURE 6 C); except for segment XI ( Fig. 6 View FIGURE 6 D), bt2 with one dorsal pair of swimming tufts of long setae (55–75 µm long) (sw t) probably formed by elongation of some round cuticular expansions; on each side of these segments three distinct lateral tubercles (lt1, lt2, lt3) ( Fig. 6 View FIGURE 6 D); two slight transversal integument folds, not extended laterally, dividing ventral surface of each segment into three welts; medium and posterior welts supporting three ventral tubercles (vt1–vt3) positioned as on Figure 6 View FIGURE 6 E; each ventral tubercle easily distinguishable by triangular cuticular expansions and with one or two sensilla ( Fig. 6 View FIGURE 6 E, F).
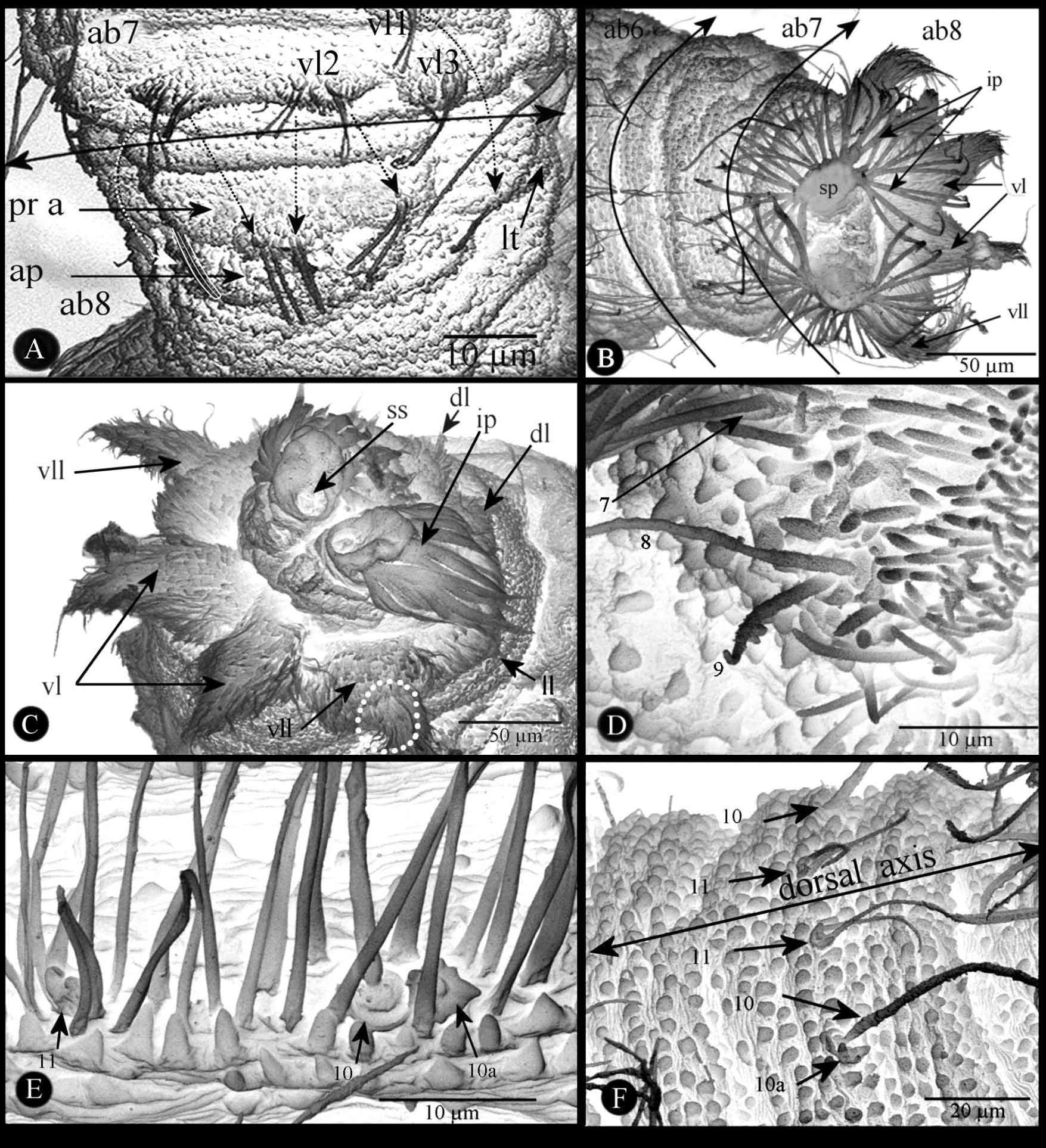
Segment XII, the last abdominal segment (ab8) is elongated ( Fig. 7 View FIGURE 7 A–D). Ventral surface with a median prominent spinulete pre-anal lobe apparently formed by fusion and stretching of the two ventral tubercles vt2 bearing stout and straight sensilla ( Fig. 7 View FIGURE 7 A, pr a) as explained below in the sensorial section. Lateral tubercle (lt) formed from vl1. Sclerotized anal plate (ap) transversally extended below the pre-anal lobe. Posterior spiracular disc with four pairs of peri-spherical lobes, entirely covered by long cuticular setae ( Fig. 7 View FIGURE 7 B, C): ventral pair (vl) conical, elongated; ventrolateral pair (vll) bisegmented, proximal part wider than elongated and conical distal portion; lateral (ll) and dorsal pairs (dl) slightly prominent. Spiracular disc with two stigmatic plates ( Fig. 7 View FIGURE 7 B, sp). Each stigmatic plate with four palmate tufts of well developed, reflecting, interspiracular processes (ip) (synonyms: float hairs or hydrofuge setae from various authors); opening shape of the stigmatic glands varying according to the stadium; stigmatic scar ( Fig. 7 View FIGURE 7 C, ss) visible only on second- and third-instar, much large and more sclerotized in the last instar.
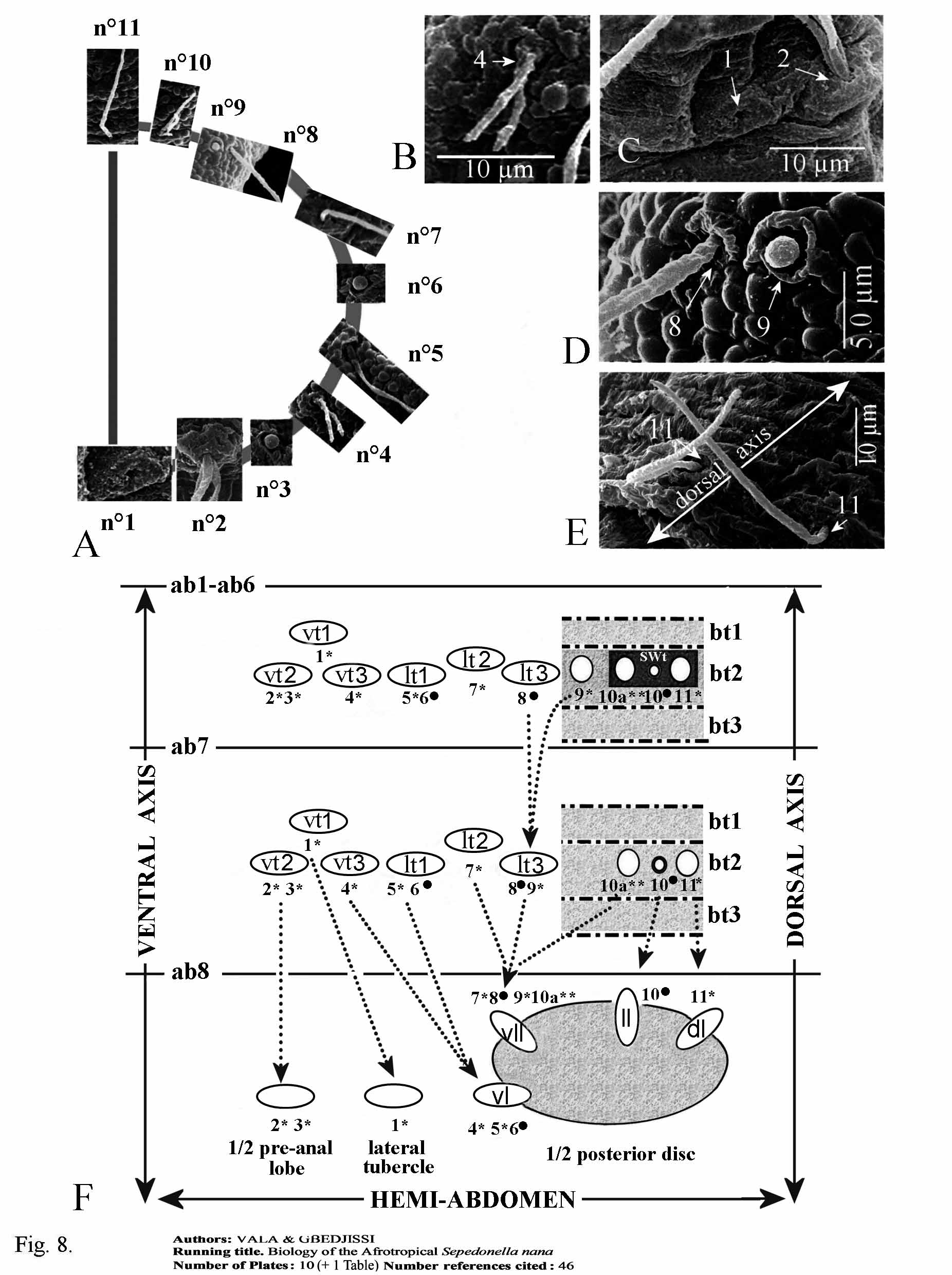
2. Sensorial receptors. The pattern of the different types of sensoria shows a symmetrical distribution along the sagittal plan of the larvae. On the side of each thoracic and abdominal segment eleven sensilla can be recognized from the ventral to the dorsal surface. These sensilla are successively numbered from 1 to 11, and addition of the letter “a” for one supplementary sensillum. The study and distribution were based on 4 specimens observed under the MEB as established by Vala (1989).
a). Cephalic segment ( Fig. 5 View FIGURE 5 E). On each cephalic lobe: one labial sensillum (ls) (coeloconicum type) on each lateral part of the mouth. One maxillary organ (mo) on dorsal surface supporting at least two very low styloconicum sensilla, unfortunately always more or less distinct. One antennal organ (ao) on the apex.
b). Thoracic segments ( Fig. 5 View FIGURE 5 D, F, G, 8A–E). One coeloconicum sensillum (n°1) inside a deep cavity, only the very small hole visible ( Fig. 8 View FIGURE 8 A, C), for this reason can be referred to the ampulaceum type. One Keilin organ (n°2) characterized by three long chaeticum sensilla (50–70 µm long) arising from a single hole ( Fig. 5 View FIGURE 5 G, 8A, C). Three basiconicum sensilla (n°3, 6, 9) (4.5–5 µm diameter), club-like ( Fig. 8 View FIGURE 8 A, D). Two chaeticum sensilla (n°4, 10), bifurcated base (10–12 µm long) ( Fig. 8 View FIGURE 8 A, B). Four long trichodeum sensilla (n°5, 7, 8, 11) (45–55 µm long) arising from a thin cuticular membranous base, often chaetum-seta broken ( Fig. 8 View FIGURE 8 A, D, E).
c). Abdominal segments ( Fig. 6 View FIGURE 6 D–F, 7, 8F). If eleven sensilla are also present on side of each segment, it is important to distinguish three abdominal portions, always from the ventral to dorsal surface. a) Segments Ab1 to Ab6. On vt1 one coeloconicum sensillum (n° 1), basal edge with two long denticulate seta-like (15–20 µm long) and doubled exteriorly by a ring of four lower expansions (5–6 µm) ( Fig. 6 View FIGURE 6 E); two similar sensilla on vt2 (n°2, 3) and one on vt3 (n°4); laterally ( Fig. 6 View FIGURE 6 D), on lt1 one coeloconicum sensillum (n°5) (seta-like, 12.5–20 µm long) plus one very long trichodeum sensillum (n°6) (55–65 µm long); on lt2 one similar sensillum (n°7) as n°5; lt3 with one isolated long trichodeum sensillum (n°8) as n°6; on latero-dorsal position one coeloconicum sensillum as the previous cited but the two setae are much longer (45–65 µm long) (n°9); dorsally, three sensilla within each tuft of cuticular expansions ( Fig. 7 View FIGURE 7 E): one flower-like coeloconicum (4–5 µm diameter) (n°10a), edge with a few short expansions (3–5 µm long); one long trichodeum sensillum (50–60 µm long) (n°10) similar to n°9; near longitudinal dorsal axis one coeloconicum (n°11) with two long seta-like expansions (55–70 µm long) on basal edge. b) Segment Abd7 ( Fig. 6 View FIGURE 6 D, 7F) sensorial equipment similar to six previous abdominal segments, except two differences: firstly, as cuticular tufts absent, sensilla n°10a, n°10, and n°11 very visible; secondly, lt3 sensillum n°9 close to n°8 c) Segment Ab8: ventrally, two pairs of stout and straight coelonicum sensilla (n°2, 3) on the pre-anal lobe ( Fig.7 View FIGURE 7 A); lateral tubercle with the vl1 sensillum n°1 after postero-lateral movement above cited; ventral lobe (vl) with sensillum (n°4) together with sensilla n°5 +6; ventro-lateral lobe (vll) with sensilla n°7–9 and 10a ( Fig.7 View FIGURE 7 D, 8F) −assemblage corroborated by Coremacera catenata [Loew, 1847] and C. marginata [Fabricius, 1775]) where lobes are not covered by long cuticular setae showing easily theses sensilla ( Vala 1989 and unpublished photos); lateral lobe (ll) and dorsal lobe (dl) with sensilla n°10 and n°11 respectively on their apex. Hypothetical movement and disposition of all abdominal sensilla are summarised on Figure 8 View FIGURE 8 F. Note: the apparent sensillum structure between n° 2 and 3 ( Fig. 6 View FIGURE 6 F) is an artefact because it was seen only once, but never on ventral surface of other abdominal segments.
As noted previously for other Sciomyzidae species ( Vala (1986, 1989) and Knutson & Vala (2011) the shape of sensilla and their setting on the segments allow differentiation of each tagma. However, for the first time we suggest an explanation concerning their distribution on the last abdominal segment (Ab8), which is distinct from the other segments by the presence of peripherical lobes. On the preanal lobe are sensilla resulting from coalescence of both ventral tubercles n°2 and 3; on lateral tubercle is the sensillum resulting from ventral tubercle n°1. Other sensilla are dispatched on the four lobes placed around the peripherical disc. Particularly, each ventrolateral lobe supports the maximum of sensilla, four here. In species having five peripherical lobes, additional dorsolateral lobes each bears one sensillum and ventrolateral lobe only three. Also, can be noted a rearrangement on the segment Ab7 where sensillum n° 9 gets closer to sensillum n° 8. In any case, from actual results, the number of 11 pair’s sensilla seems to be constant on each thoracic and abdominal segment for all studied species of sciomyzid larvae.
d). Cephalopharyngeal skeleton. All characteristics are described along with the morphology of each individual instar larva
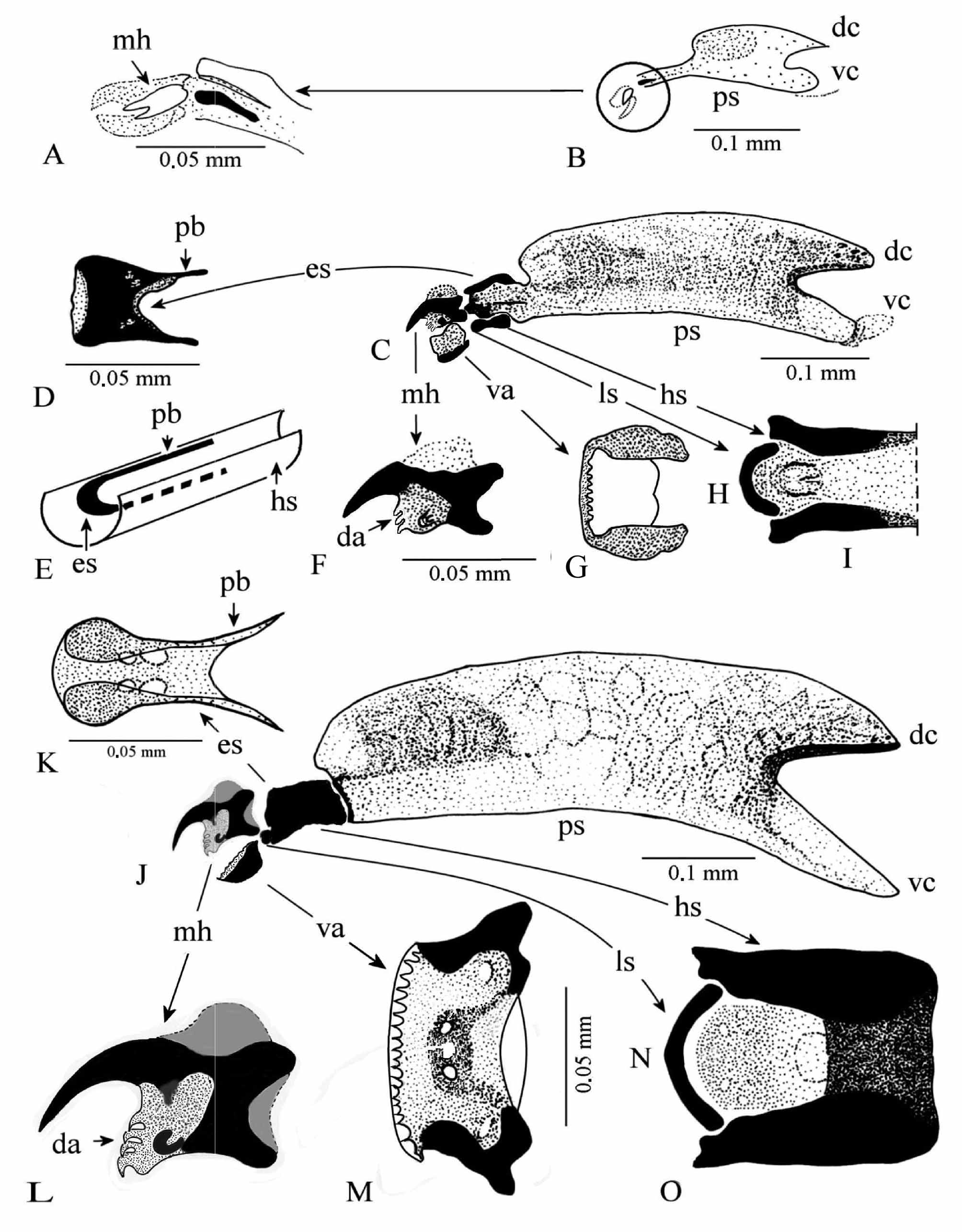
First instar larva (L1) ( Fig. 5 View FIGURE 5 D–G; 6C–F; 7A, B; 9A, B). Length 0.81–1.23 mm, greatest width 0.18–0.23 mm. Transparent aspect. Anterior spiracles absent. Four palmate interspiracular processes ( Fig. 7 View FIGURE 7 B, ip) 50.12–53.05 μm long. Posterior spiracular opening B-shaped. Cephalopharyngeal skeleton ( Fig. 9 View FIGURE 9 A, B). 0.19–0.23 mm long, light yellow-brownish. Mouthhook (mh) brown, anteriorly pointed, accessory teeth absent. Ventral arch tiny, 8–10 anterior denticles. Hypostomal sclerite very light except brown longitudinal axe. Ligulate sclerite very clear, barely visible. Epistomal sclerite light brown, above hypostomal sclerite as parastomal bars short, and epistomal sclerite. Paired pharyngeal sclerite (ps) lightly pigmented, joined along ventral margins, dorsal cornua (dc) with pointed apex, ventral cornua (vc) apex large, both cornua with hyaline processes and without apparent windows. (Based on 10 specimens).
Second instar larva (L2) ( Fig. 6 View FIGURE 6 A; 7C–F; 9C–I). Length 2.51–3.20 mm; greatest width 0.50–0.80 mm. General coloration greyish to dark brown in young L2 then light grey and 3 motley longitudinal zones on dorsal surface. Anterior spiracles (as) small, 3–5 µm long, delimited by a thin membrane (mb), three more or less lobulated and flattened papillae ( Fig. 6 View FIGURE 6 A, pa). Each spiracular plate (sp) with one spiracular scar (ss) with sclerotized edge, three oval spiracular openings and four thick interspiracular processes (ip) (Fig, 7C).
Cephalopharyngeal skeleton ( Fig. 9 View FIGURE 9 C–I) hardly sclerotized. Length 0.42–0.48 mm. Brown-yellowish. Mouthhook (mh), 0.05 mm length, hook very pointed with 3 incurved accessory teeth (da) below hook, one small foramen in the middle, one strong posterodorsal and one strong posteroventral extension. Ventral arch (va) bilobed, anterior margin turned up with 10–14 denticles posteriorly directed, lateral sides turned up. Epistomal sclerite (es) with lateral sides curved down. Parastomal bars very strong (pb). Ligulate sclerite (ls) arc shaped. Hypostomal sclerite bilobed, lateral sides turn up ( Fig. 9 View FIGURE 9 E, G, hs). Pharyngeal sclerite, 0.31 mm length ( Fig. 9 View FIGURE 9 C, ps). (Based on 10 specimens).
Third instar larva (L3) ( Fig. 6 View FIGURE 6 B; 9J– O; 10A–B). Length 4.00–6.00 mm; greatest width 1.00– 1.70 mm. Uniformly greyish or very motley, dorsally five transversal stripes alternatively dark and pale. Anterior spiracle, elongated, 8–10 µm long, five papillae, base with one round stigmatic scare (ss) ( Fig. 6 View FIGURE 6 B).
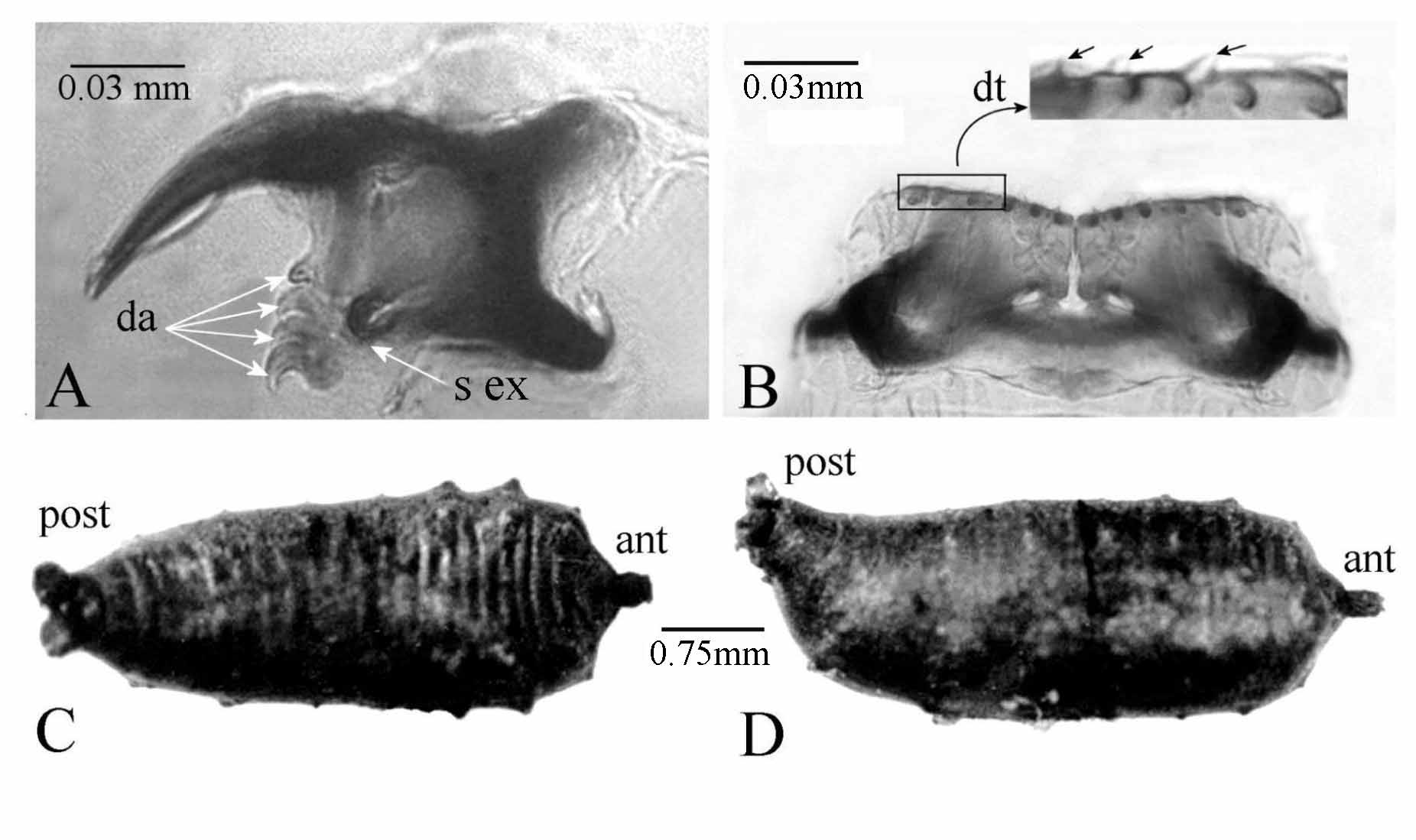
Cephalopharyngeal skeleton ( Fig. 9 View FIGURE 9 J– O; 10A, B), very dark, strongly sclerotized, less pigmented areas developing on wing-like pharyngeal sclerites. Median length 0.61 ± 0.10 mm. Epistomal sclerite (es) and parastomal bars (pb) fused ( Fig. 9 View FIGURE 9 K), strongly pigmented. Mouthhook 0.08 mm, hook very incurved, four accessory teeth (9L, 10A, da), presence of one strongly sclerotized expansion behind the accessory teeth ( Fig. 10 View FIGURE 10 A, s ex), one foramen well visible. Ventral arch (va) bilobed, lateral sides up turned, many tiny denticles (dt) on anterior margin ( Fig. 9 View FIGURE 9 M; 10B, dt). Lingual sclerite (ls) arc-shape located between anterior arms of hypostomal sclerite ( Fig. 9 View FIGURE 9 N). Hypostomal sclerite (hs), both sides enveloping the complex epistomal sclerite and parastomal bars ( Fig. 9 View FIGURE 9 O). Pharyngeal sclerite (ps) length 0.40 mm, massive, intense pigmentation, dorsal cornua (dc) shorter than ventral cornua (vc), both without window ( Fig. 9 View FIGURE 9 J). Indentation index 0.18. (Based on 14 specimens).
Puparium ( Fig. 10 View FIGURE 10 C, D). Length 3.40–4.70 mm, greatest width 1.20–1.90 mm. Brown to black. Dorsal surface ( Fig. 10 View FIGURE 10 C) with two brown transversal stripes separated by one whitish transversal stripe. Ventral surface uniformly brownish to dark. Dorsal cephalic cap bearing conspicuous anterior spiracles at anterolateral corners. Dorsal and lateral tubercles, round, slowly expanded, well visible mostly in anterior part ( Fig. 10 View FIGURE 10 C). Anterior end (ant) in prolongation to median longitudinal axis ( Fig. 10 View FIGURE 10 D). Posterior end (post) over axis and turned up, distinctly divergent spiracular tubes. Ventral and ventrolateral lobes withered below constricted disc; interspiracular processes vestiges of L3 flattened ( Fig. 10 View FIGURE 10 D). Anal plate black. (Based on 8 specimens).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
SubClass |
Oligochaeta |
|
Order |
|
|
Family |
|
|
Genus |
Sepedonella nana
| Vala, Jeanclaude & Gbedjissi, Louis Ghélus 2011 |
Sepedon lippensi
| Verbeke 1950 |
S. umbrosa
| Verbeke 1950 |
S. ornatifrons
| Adams 1910 |