
Campydoroides manautei Holovachov, 2019
|
publication ID |
https://doi.org/ 10.5852/ejt.2019.518 |
|
publication LSID |
urn:lsid:zoobank.org:pub:7DDF6B2E-BC13-459E-A147-6C2ADD43185C |
|
DOI |
https://doi.org/10.5281/zenodo.5613679 |
|
persistent identifier |
https://treatment.plazi.org/id/CD624B5D-3F24-4D54-9C3C-C00615BB340E |
|
taxon LSID |
lsid:zoobank.org:act:CD624B5D-3F24-4D54-9C3C-C00615BB340E |
|
treatment provided by |
Plazi |
|
scientific name |
Campydoroides manautei Holovachov |
| status |
gen. et sp. nov. |
Campydoroides manautei Holovachov gen. et sp. nov.
urn:lsid:zoobank.org:act:CD624B5D-3F24-4D54-9C3C-C00615BB340E
Figs 2–4 View Fig View Fig View Fig ; Table 3 View Table 3
Diagnosis
Campydoroides manautei sp. nov. is characterised by a 0.49–0.82 mm long body, papilliform inner labial, outer labial and cephalic sensilla, amphidial opening a transverse slit, strongly cuticularised stoma with single robust dorsal tooth, muscular pharynx with distinct basal bulb, large cardia, didelphic, amphidelphic female gonads, straight vagina, and elongate conoid tail equal to 6.0–9.6 anal body diameters in length.
Etymology
The new species is named in honour of Joseph Manauté, director of the Parc provincial de la Rivière bleue, for his indispensable support during the sampling in the Park.
Type material
Holotype NEW CALEDONIA • 1 ♀; Yate Province , Parc provincial de la Rivière bleue, station HYNC2372 /4680; 22°04.401′ S, 166°36.995′ E; 18 Jul. 2018; O. Holovachov leg.; virgin forest, soil; MNHN-BN511 . GoogleMaps
Paratypes
NEW CALEDONIA • 4 ♀♀; Yate Province , Parc provincial de la Rivière bleue, station HYNC2372 /4680; 22°04.401′ S, 166°36.995′ E; 19 Nov. 2017 and 18 Jul. 2018; O. Holovachov leg.; virgin forest, soil; MNHN-BN511 , SMNH Type-9167 GoogleMaps • 2 ♀♀; Yate Province , Parc provincial de la Rivière bleue, station HYNC2378 ; 22°05.374′ S, 166°38.185′ E; 19 Nov. 2017; O. Holovachov leg.; virgin forest, wet soil at the bottom of a stream bed; SMNH Type-9168, SMNH Type-9169 GoogleMaps .
Additional non-type material NEW ZEALAND • 2 ♀♀; Napier, Puketitiri, station Z236; 10 Nov. 1988; D. Sturhan leg.; SMNH- 175283 .
Description
Female
Body cylindrical, tapering slightly along pharyngeal region towards truncated anterior end, tapering more rapidly along tail towards tail tip; straight or ventrally curved upon fixation. Cuticle finely transversely striated along entire body, starting from base of outer labial sensilla and ending near tail tip; striations less than 1 µm; lateral alae absent; any other types of cuticular ornamentation absent. Body pores and epidermal glands absent. Somatic sensilla distinct along anteriormost part of pharyngeal region (cervical setae, see below) and on tail, indistinct or absent along rest of body. Labial region truncated, lips indistinct (fused). Six inner labial sensilla small papilliform, located on anterior surface of lips. Six outer labial sensilla small papilliform, located on outer periphery of labial region, 1.5–5.0 µm from anterior end (5 µm in specimen with everted stoma). Four cephalic sensilla small papilliform, located short distance behind lip region, 4–7 µm from anterior end, at level with dorsal tooth tip (in relaxed specimens, with inverted stoma, see below). Four pairs of cervical sensilla present, papilliform in shape, arranged in one dorsosublateral, one subdorsal and two subventral pairs at level of stoma base. Amphid stirrup-shaped with transverse slit-like opening 3–4 µm wide, located at level with middle of stoma. Oral opening
apical, surrounded by a fringe of tiny projections, visible only when stoma is everted to the outside ( Fig. 2A View Fig ). Stoma conoid; cheilostom short and weakly cuticularised; gymnostom as long and broad as cheilostom, weakly cuticularised; stegostom with strongly cuticularised walls, distinctly subdivided into three sections by breaks in cuticularised walls, with large dorsal tooth projecting anteriad from its middle section. Tooth heavily cuticularised, 3–5 µm long, hollow inside, protrusible, its tip is 10–13 µm from stoma base. Pharyngeal tubes absent. Pharynx cylindrical anteriorly, with distinct basal bulb; anterior cylindrical part of pharynx with evenly distributed myofilaments, basal bulb strongly muscularised; pharyngeal lumen uniform in thickness along entire pharynx length; valves absent. Pharyngeal glands and their orifices indistinct. Cardia large, ovoid, partially embedded into intestinal tissue. Nerve ring encircling pharynx at 43–54% of its length. Secretory-excretory system indistinct/absent. Female reproductive system didelphic, amphidelphic, ovary branches reflexed antidromously, usually located on same side of intestine; symmetrical in younger females, asymmetrical in older females when large developing ovocyte or intrauterine egg is present in one of genital tubes. Anterior genital tube 21–91 µm long, equal to 4–13% of total body length, situated to either right (n = 4) or left (n = 3) of intestine; posterior genital tube 23–55 µm long, equal to 4–9% of total body length, situated to either right (n = 3) or left (n = 4) of intestine. Vulva a transverse slit, located anterior to midbody. Vagina straight, equal to 0.2–0.3 body diameters in length, with developed sphincter muscle surrounding its proximal part; pars refringens vaginae or epiptygmata absent. Spermatheca absent. Intrauterine egg measuring 64–77 × 17– 21 µm. Rectum short, equal to 0.8–1.2 anal body diameters in length. Tail elongate, distinctly subdivided into conoid proximal part 26–42 µm in length, and subcylindrical distal part 76–136 µm long. Caudal setae present, one subdorsal pair along proximal conoid part of tail, one subventral pair at level with junction between proximal and distal parts of tail, and single lateral setae on left side of distal cylindrical part of tail. Caudal glands present, opening via small terminal spinneret.
Male
Not found. The species is presumed to be thelytokous.
Remarks
The two female specimens collected in New Zealand match very well the morphology of the type specimens from New Caledonia, except for the larger body size, higher values for some body measurements and longer tail (including higher c’ -ratio). Since no qualitative morphological differences were found between the two populations, they both are here considered to belong to the same species.
Notes on the systematics of Campydorina
The unique and poorly understood morphology of the genus Campydora hindered understanding of its relationships with other nematodes and resulted in the taxonomic inflation from the genus to the suborder level ( Jairajpuri 1983). This decision was followed in subsequent publications, although the suborder was transferred from the order Dorylaimida to the order Enoplida based on the morphological and molecular evidence, and its relationships within the order Enoplida remained unresolved ( Siddiqi 1983; Winiszewska 2001; Mullin et al. 2003; Zhao et al. 2012; Smol et al. 2014).Subsequent phylogenetic studies using 18S rDNA sequence suggested close affinities between the genera Campydora , Rhabdolaimus de Man, 1880 and Syringolaimus de Man, 1888 , placing these three genera as a monophyletic clade within the unresolved order Enoplida ( Meldal et al. 2007; van Megen et al. 2009; Bik et al. 2010; Shokoohi et al. 2013; Smythe 2015; Leduc et al. 2018). This grouping can be further substantiated by at least one morphological character: the presence of a well-developed basal bulb, not found in any other Enoplida (the genera Polygastrophora de Man, 1922 and Belbolla Cobb, 1920 have the posterior part of the pharynx modified into a series of bulb-like swellings that are of independent origin). Placement of the genus Syringolaimus in the suborder Campydorina is further supported by the monorchic male reproductive system similar to the reproductive system found in the species from the genera Campydora and Rogerus ( Hoeppli & Chu 1932; Winiszewska 2001; Tchesunov 2017), while it is diorchic in other genera of Ironina Siddiqi, 1983 ( Shokoohi et al. 2013; Chen & Guo 2015). The revised classification of the suborder Campydorina is given below.
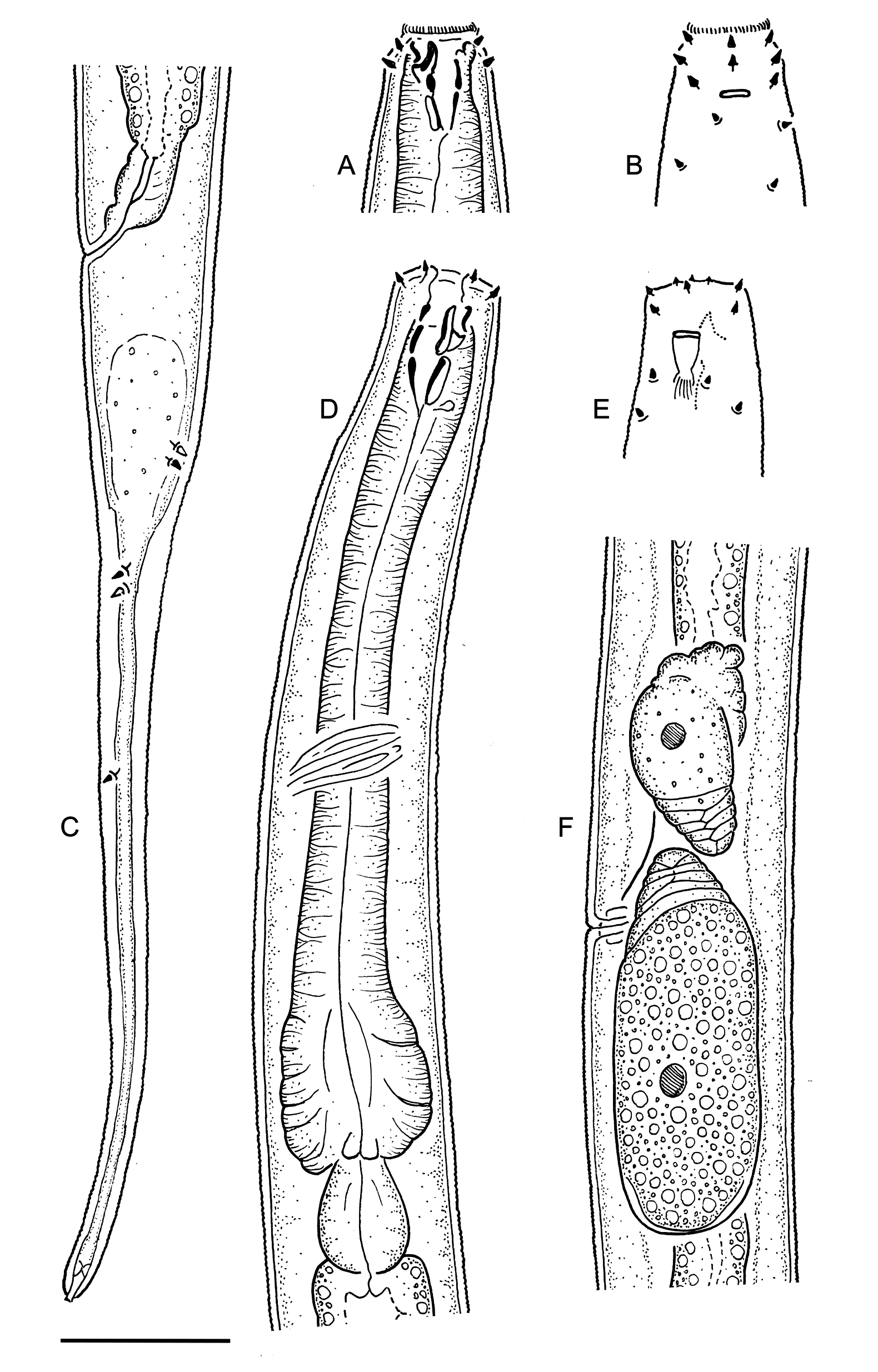
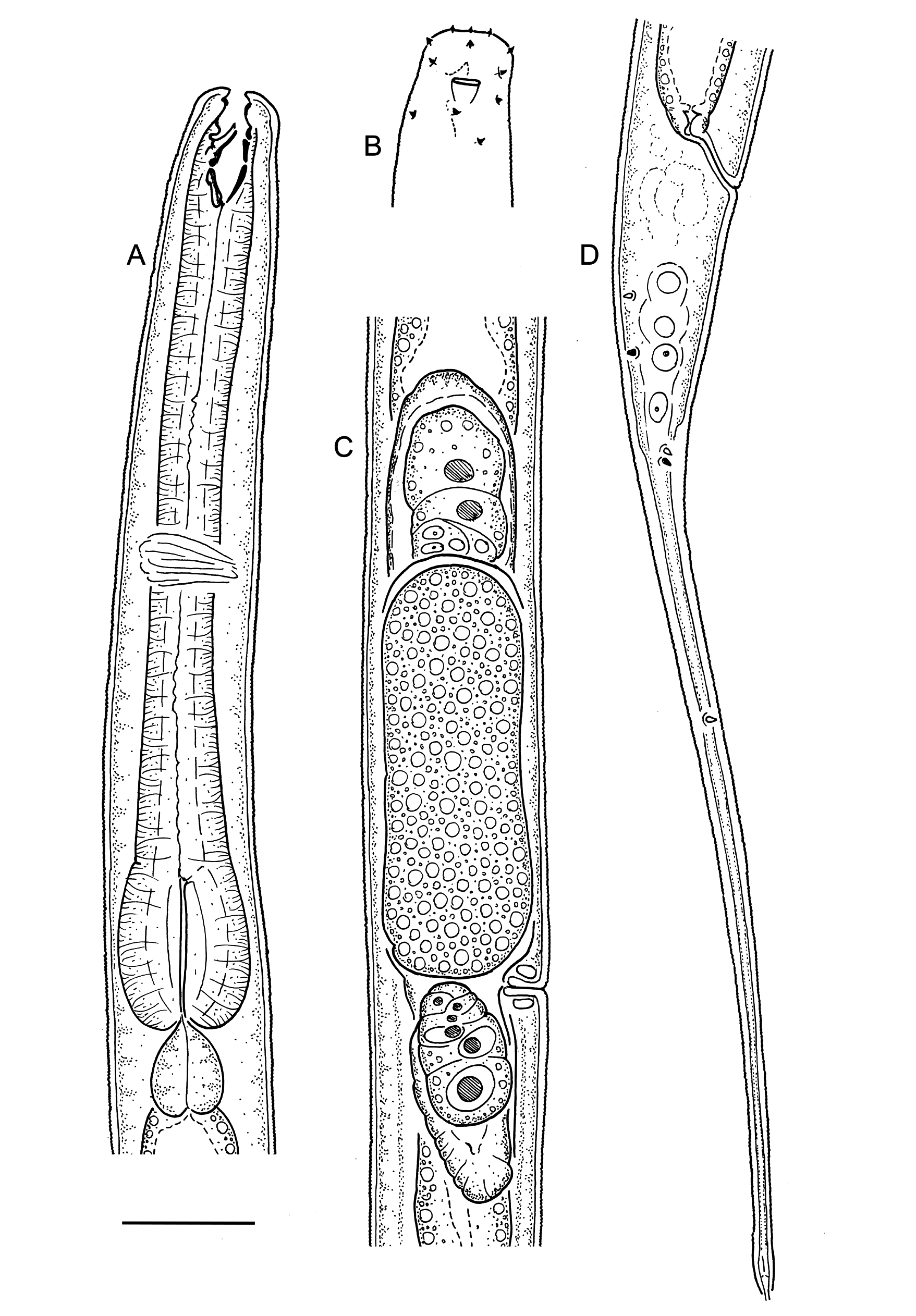
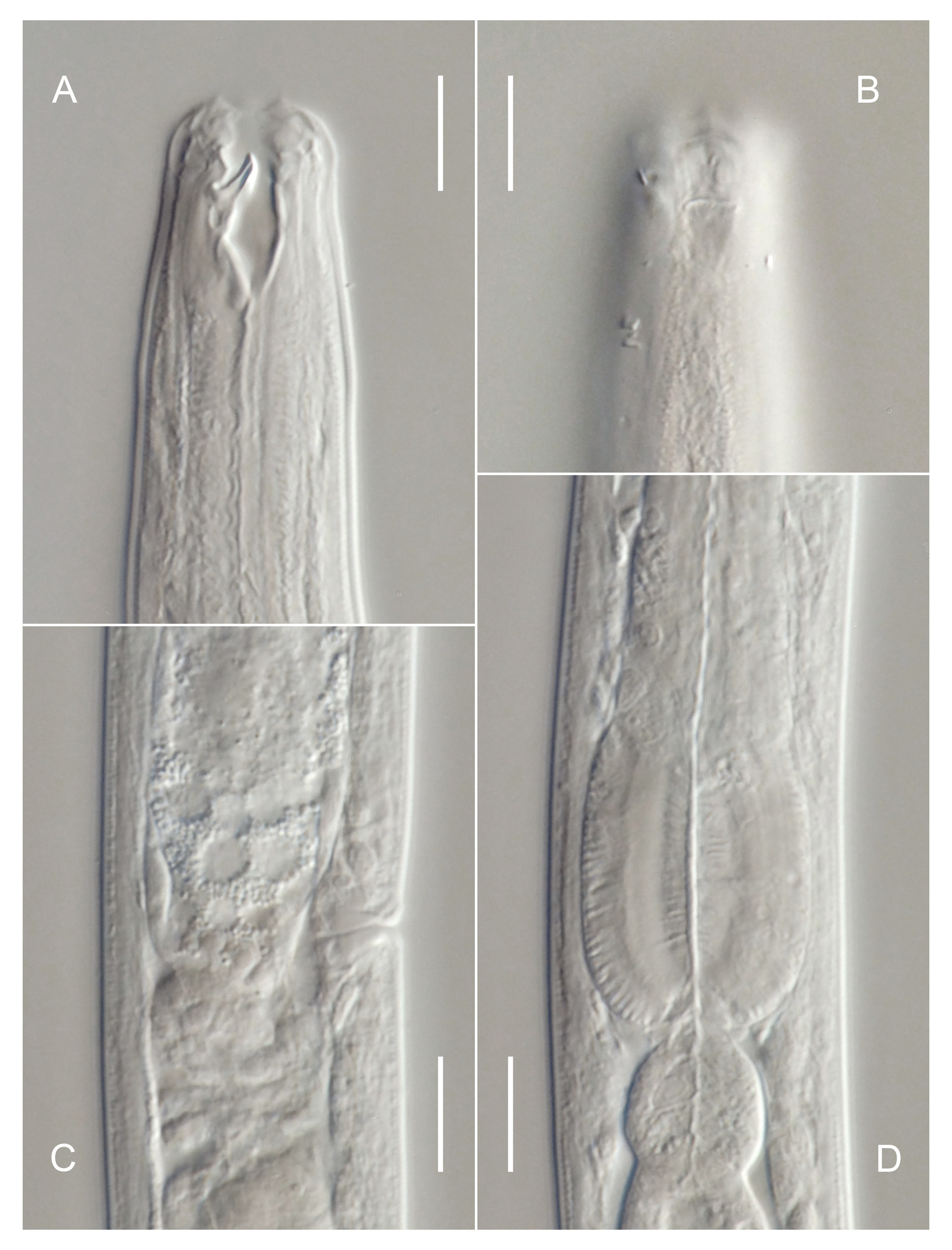
Table 3. Morphometrics of Campydoroides manautei Holovachov gen. et sp. nov. (all measurements are in µm, except for the ratios a, b, c, c’ and V).
| Character | Holotype female | New Caledonia (n = 7, incl. holotype) | New Zealand (n = 2) |
|---|---|---|---|
| Body length | 609 | 537 ± 45 (492–609) | 786; 817 |
| Body diameter | 26 | 24 ± 3 (19–28) | 25; 28 |
| Pharynx length | 110 | 94 ± 9 (86–110) | 140; 146 |
| Tail length | 128 | 115 ± 9 (104–128) | 173; 178 |
| Anal body diameter | 16 | 16 ± 1 (15–17) | 18; 19 |
| a | 23.9 | 22.7 ± 2 (19.5–25.5) | 31.6; 29.6 |
| b | 5.6 | 5.7 ± 0.4 (4.9–6.1) | 5.6; 5.6 |
| c | 4.8 | 4.7 ± 0.2 (4.3–5.0) | 4.6; 4.6 |
| c’ | 8.0 | 7.1 ± 0.8 (6.0–8.0) | 9.6; 9.2 |
| V (%) | 40.5 | 41.4 ± 1.1 (40.5–43.4) | 42.1; 42.6 |
| Labial region diameter | 12 | 12 (12–14) | 14; 14 |
| Amphid from anterior end | 7 | 8 (7–9) | 9; 8 |
| Stoma length | 17 | 16 (13–17) | 19; 17 |
| Stoma width | 6 | 6 (5–6) | 6; 6 |
| Nerve ring from anterior end | 59 | 47 ± 7 (37–59) | 75; 76 |
| Basal bulb length | 20 | 21 ± 2 (17–23) | 26; 23 |
| Basal bulb width | 19 | 17 ± 1 (16–19) | 20; 21 |
| Cardia length | 14 | 9 ± 3 (6–14) | 14; 15 |
| Cardia width | 10 | 11 ± 1 (10–12) | 12; 12 |
| Vagina length | 7 | 6 (6–7) | 7; 7 |
| Rectum length | 15 | 14 ± 1 (12–16) | 15; 15 |
| Cardia width | 10 | 11 ± 1 (10–12) | 12; 12 |
| Vagina length | 7 | 6 (6–7) | 7; 7 |
| Rectum length | 15 | 14 ± 1 (12–16) | 15; 15 |
| SMNH |
Department of Paleozoology, Swedish Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |