
Nucinella boucheti, Perna, Rafael La, 2005
|
publication ID |
https://doi.org/ 10.5281/zenodo.273223 |
|
DOI |
https://doi.org/10.5281/zenodo.6264975 |
|
persistent identifier |
https://treatment.plazi.org/id/03930553-FFCE-3018-FEC1-FC111EE2FE4D |
|
treatment provided by |
Plazi |
|
scientific name |
Nucinella boucheti |
| status |
sp. nov. |
Nucinella boucheti n.sp.
Figs. 2 View FIGURE 2 , 3 View FIGURE 3
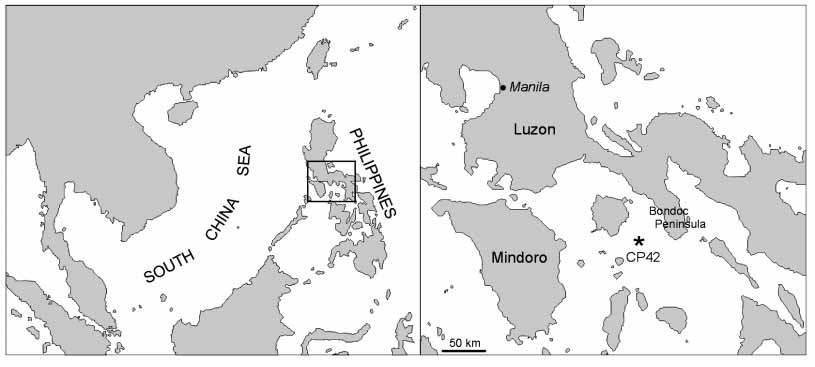
Type locality. MUSORSTOM 2 st. CP 42, 13 °05’N– 122°25’E, 1610– 1580 m (25.11.1980), southwest off Bondoc Peninsula, Luzon Island, Philippines ( Fig. 1 View FIGURE 1 ).
Type material. Holotype (right valve) and 18 paratypes ( MNHN, 4411, 4412).
Material examined. Only the type material, consisting of 18 valves and a single livecollected specimen. The valves’ surface is somewhat corroded, particularly on the umbo, and the periostracum is mostly lost.
Etymology. Named after the French malacologist Philippe Bouchet, in honor of his intense research activity on tropical West Pacific molluscs.
Distribution. Only known from the type locality.
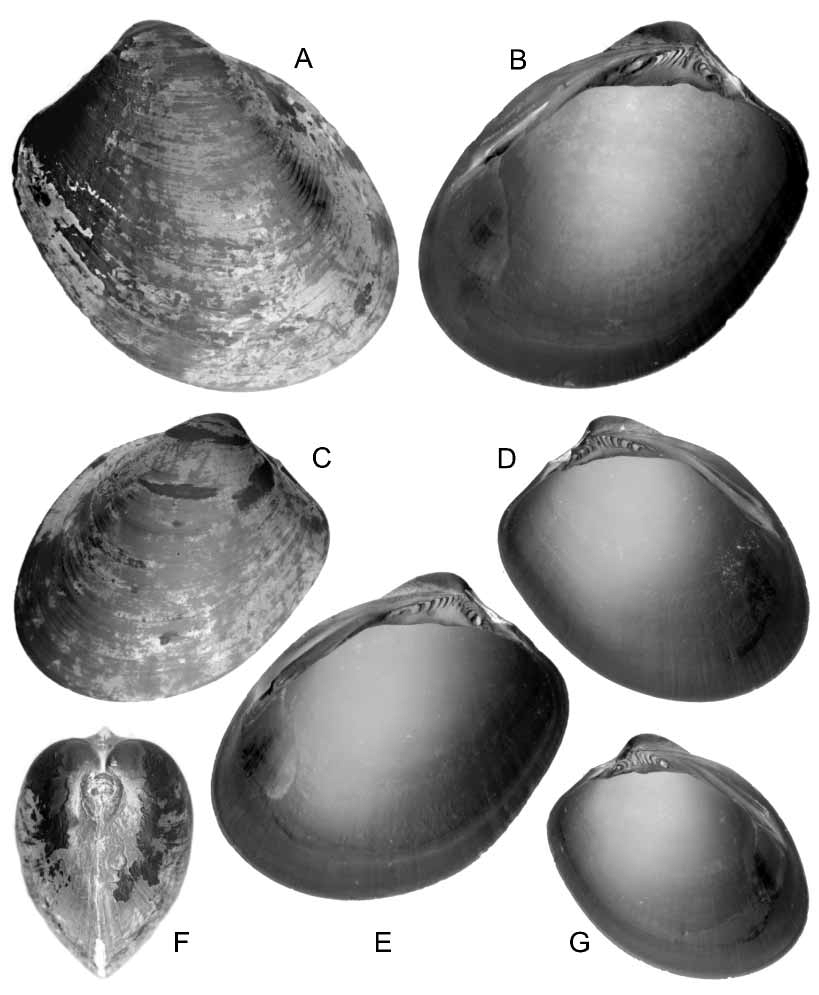
Diagnosis. An unusually large species for the family, up to 25 mm in height, with a markedly convex, nuculiform shell and a delicate hinge.
Description. Shell equivalve, strongly inequilateral, obliquely ovate, nuculiform, markedly convex. Shell wall thin, fragile. Umbo large, projecting well beyond dorsal margin, opisthogyrous. Anterodorsal margin short, convex, obtusely curving to a slightly convex anterior margin. Ventral margin strongly convex, smoothly curving to a long, barely convex posteroventral margin. Posterodorsal margin short, slightly concave, forming a smooth, narrow curve when joining to posterior margin. Junction between posterodorsal and posteroventral margin as an obtuse smooth angulation. Hinge plate not strong, wide, extending from posterodorsal margin to about 2/3 of anterior side length, with a wide, flat, somewhat triangular area below dentition. Subumbonal teeth of different shape, size and orientation, in a single, arched series, numbering about 13 in holotype. Posterior teeth mainly crescentshaped, the last one is a blunt tubercle. Below beak, few irregularly shaped to lamellar teeth. Anterior teeth mainly chevronshaped. Lateral dentition consisting of a rather strong, lamellar, grossly triangular tooth, continuous with hinge plate margin, with rather deep longitudinal grooves, and a wide submarginal socket with 2–3 small ridgelike teeth inside. Ligament pit external, wide, immediately posterior to beak, occupying all posterodorsal margin. Ligament pit wide, deep; pit margins with a deep groove for ligament insertion. Ligament thick, strongly bulging, partly covered by a thick periostracum. Inner valve surface white, dull. Pallial line fairly well distinct, no sinus. Anterior muscle scar large, ovate, marginal in position, below shell height midpoint; no posterior scar. Outer surface with shallow, ill defined, concentric riblets and thin growth striae. Periostracum rather thick, shining, varnishlike, greenishbrowinsh, mostly not preserved. Prodissoconch not seen.
Dimensions. Holotype 24 mm in height, 25 mm in length, 6 mm in thickness. Paratypes 15–25 mm in height.
Remarks. Because of its large size and distinct hinge (see below), N. boucheti is strikingly different from all the other species in the genus. The sole species which may deserve some comparisons with the present one, is N. maxima (Thiele and Jaeckel, 1931) , a deepsea species from Zanzibar Strait. This species is notably larger (12.5 mm) than the usual size range of the family and it actually was the largest known species before the present description. Under this name, Kusnetsov and Schileyko (1984) reported a deepsea species from the Gulf of Aden, but their species is much smaller (about 3 mm) and roughly rectangular in shape. Another particularly large species is N. viridis ( Matsukuma et al., 1982) (about 10 mm), from the Central East Japan in 500 m. Like the present species, N. viridis has a marked nuculiform shape, but its hinge consists of few, rather coarse teeth.
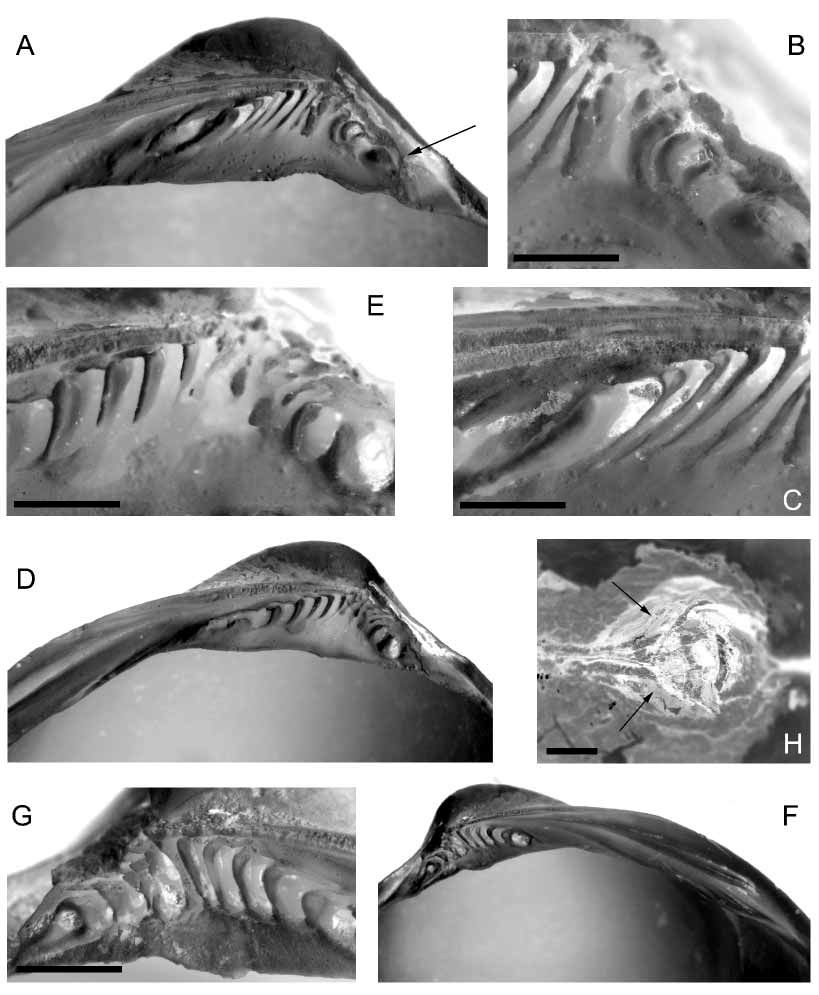
The hinge of N. boucheti contrasts with the typical hinge of the genus (Bernard 1987; Vokes 1956; Matsukuma et al. 1982; Pojeta 1988; Beu & Maxwell 1990; La Perna 2004), which has few subumbonal teeth (generally not exceeding six), somewhat different in size and orientation, rather stout (“peglike”) and coarse, giving a rather strong appearance. In spite of the large shell size, the present species has small and thin subumbonal teeth, giving a delicate appearance to the hinge, and they also exhibit some unusual shapes. The posterior teeth are oddly crescentshaped, with the convex side towards the beak ( Fig. 3 View FIGURE 3 B), whereas the last posterior one is stout, roundish to elliptical, or with a strange circular shape ( Fig. 3 View FIGURE 3 G). The central teeth are thin, lamellar or irregularly shaped (resorbed?) ( Figs. 3 View FIGURE 3 B, E, G). The anterior teeth are slightly stronger, asymmetrically chevronshaped, with a short dorsal branch parallel to shell margin ( Figs. 3 View FIGURE 3 E, C), whereas the most distal ones are elliptical or elongated ( Figs. 3 View FIGURE 3 A, C). Another difference consists in the arrangement of the subumbonal teeth, which do not extend all through the hinge plate height, whereas in the other species most part of hinge plate is occupied by teeth. The lateral dentition is similar to that of the other species, but the large lamellar tooth has longitudinal grooves ( Fig. 3 View FIGURE 3 F).
The ligament is thick, strongly bulging and partly covered by a rather thick periostracum ( Fig. 3 View FIGURE 3 H), within a wide and deep attachment area, whose margins forms a deep groove in the shell wall for the ligament insertion ( Figs. 3 View FIGURE 3 A, D). There is a thin external lamellar layer, and a thicker, partly mineralised, coarsely fibrous internal layer. The ligament attachment area of N. boucheti is not different from that of the other species in the genus (e.g., Pojeta 1988: pl. 5, fig. 3; pl. 7 fig. 6; Beu and Maxwell 1990, pl. 50, fig. b), but the large size allows the observations by Waller (1990; 1998) to be confirmed, i.e. the lack of nymphs in nucinellids (which are instead present in the other solemyoids).
| MNHN |
Museum National d'Histoire Naturelle |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
SubClass |
Protobranchia |
|
Order |
|
|
Family |
|
|
Genus |