
Nebria (Barbonebriola), Hube & Schmidt, 2017
|
publication ID |
https://doi.org/ 10.5169/seals-787049 |
|
DOI |
https://doi.org/10.5281/zenodo.6315653 |
|
persistent identifier |
https://treatment.plazi.org/id/03948799-FFD2-FFD0-FF2B-FC88FE43FEEE |
|
treatment provided by |
Carolina |
|
scientific name |
Nebria (Barbonebriola) |
| status |
subgen. nov. |
Barbonebriola subgen. nov.
Type species: N. barbata ANDREWES, 1929, p. 130 View in CoL , by present designation.
= ganeshi View in CoL species group sensu Ledoux & Roux (2005)
Remarks to the ganeshi species group sensu Ledoux & Roux (2005)
Up to now the ganeshi species group consists, in accordance with Ledoux & Roux (2005), of the three taxa N. ganeshi , N. barbata and N. restricta .
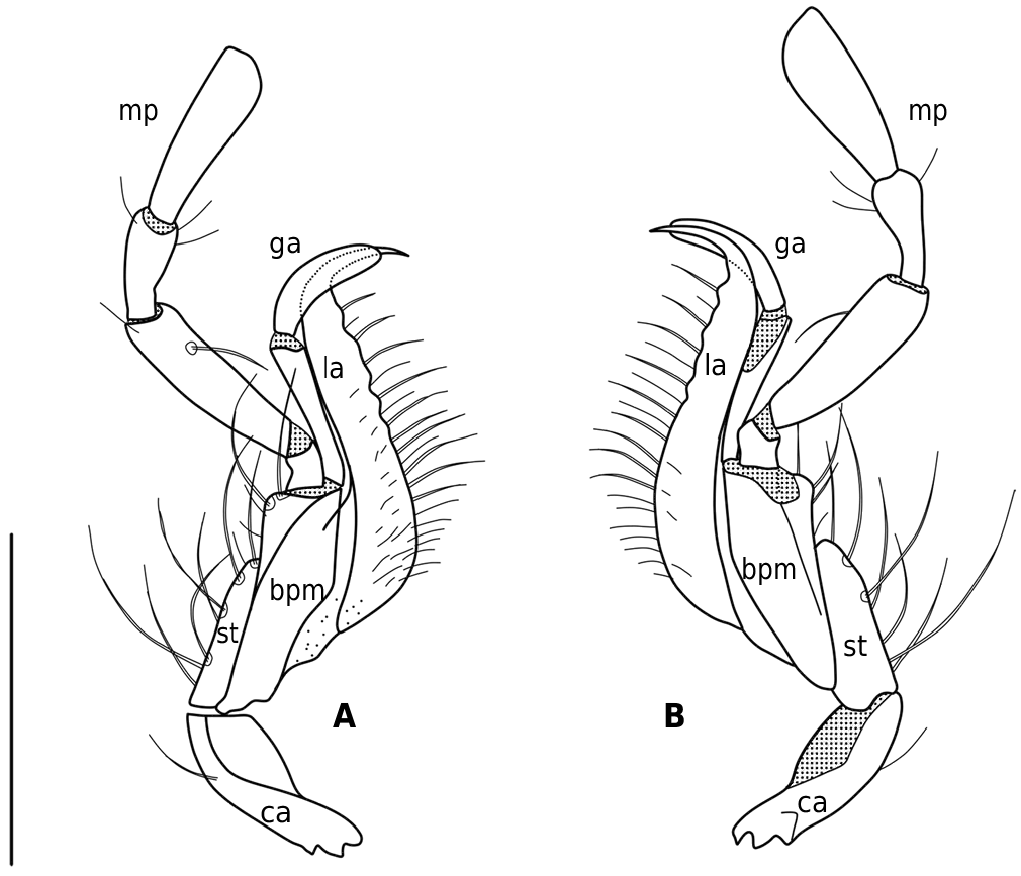
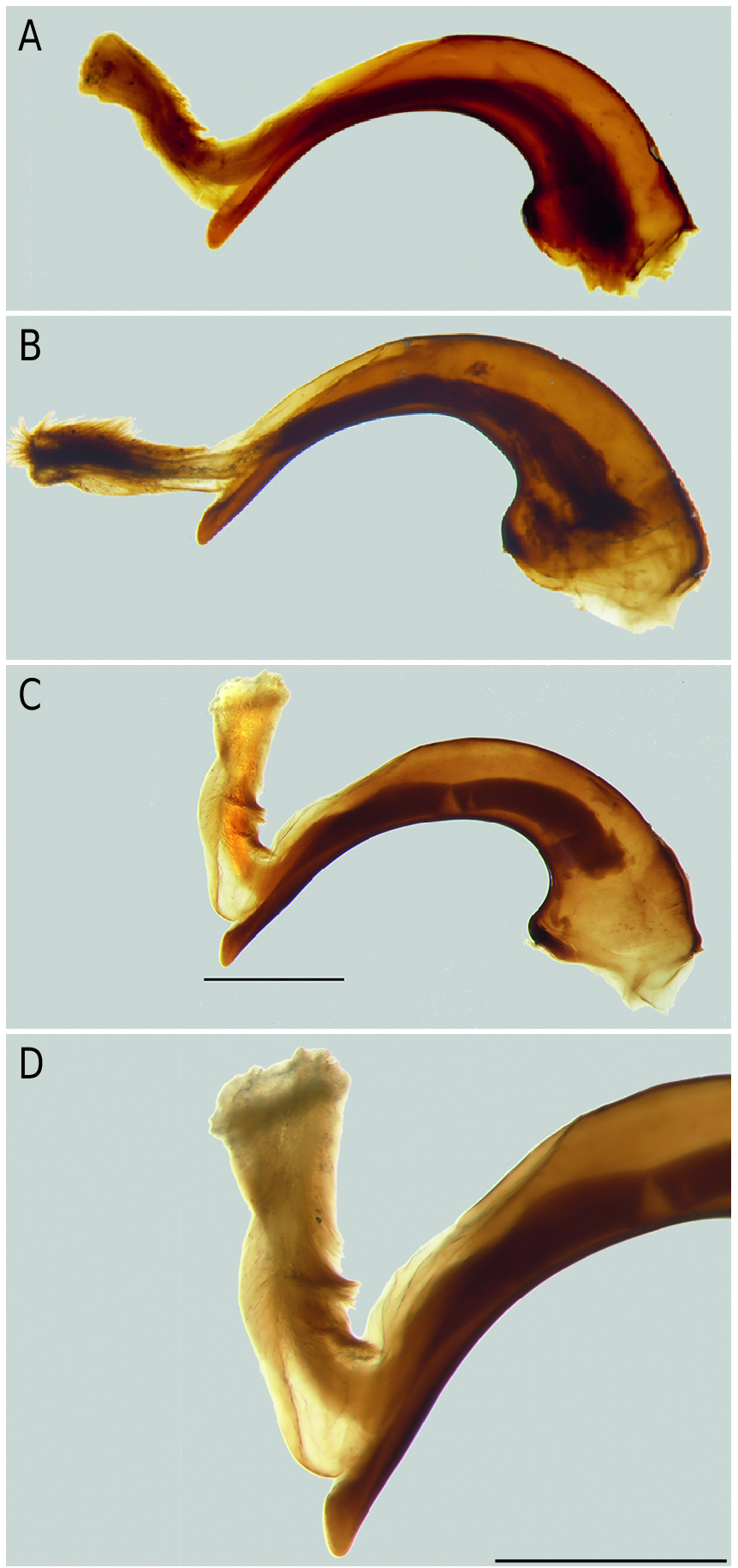
The examination of a considerable amount of specimens of the ganeshi species group (with the exception of N. restricta ) just as of three new species which are described below revealed an exclusive morphologic character common to all members of this group: The stipes as the second maxillomere is developed to a latero-external berry-like swelling bearing the setae on distinct tubercles ( Fig. 1 View Fig ). All other Epinebriola species outside of the ganeshi species group are characterized by the presence of a "normal" flattened stipes which represents the plesiomorphic state ( Fig. 2 View Fig ).
Ledoux & Roux (2005) established the ganeshi species group probably due to reasons of distribution and/or habitus resemblance, but without giving any precise morphological characters. Specimens of N. barbata (from Barbusa La near Chilas, Pakistan) and of N. ganeshi (from the type locality Rohtang La, Himachal Pradesh, India) have a conspicuous berry-like bulge on the stipes ( Fig. 1 View Fig , 3 View Fig ) as well as the new species N. rubostipes sp. nov., N. tenuisulcata sp. nov. and N. kagmara sp. nov. Such a chaetiphorous lateral bulge of the maxillary stipes has neither been noted or described so far nor used in the species identification within the subgenus Epinebriola , not even within the genus Nebria . Obviously this maxillary character has consistently been overlooked.
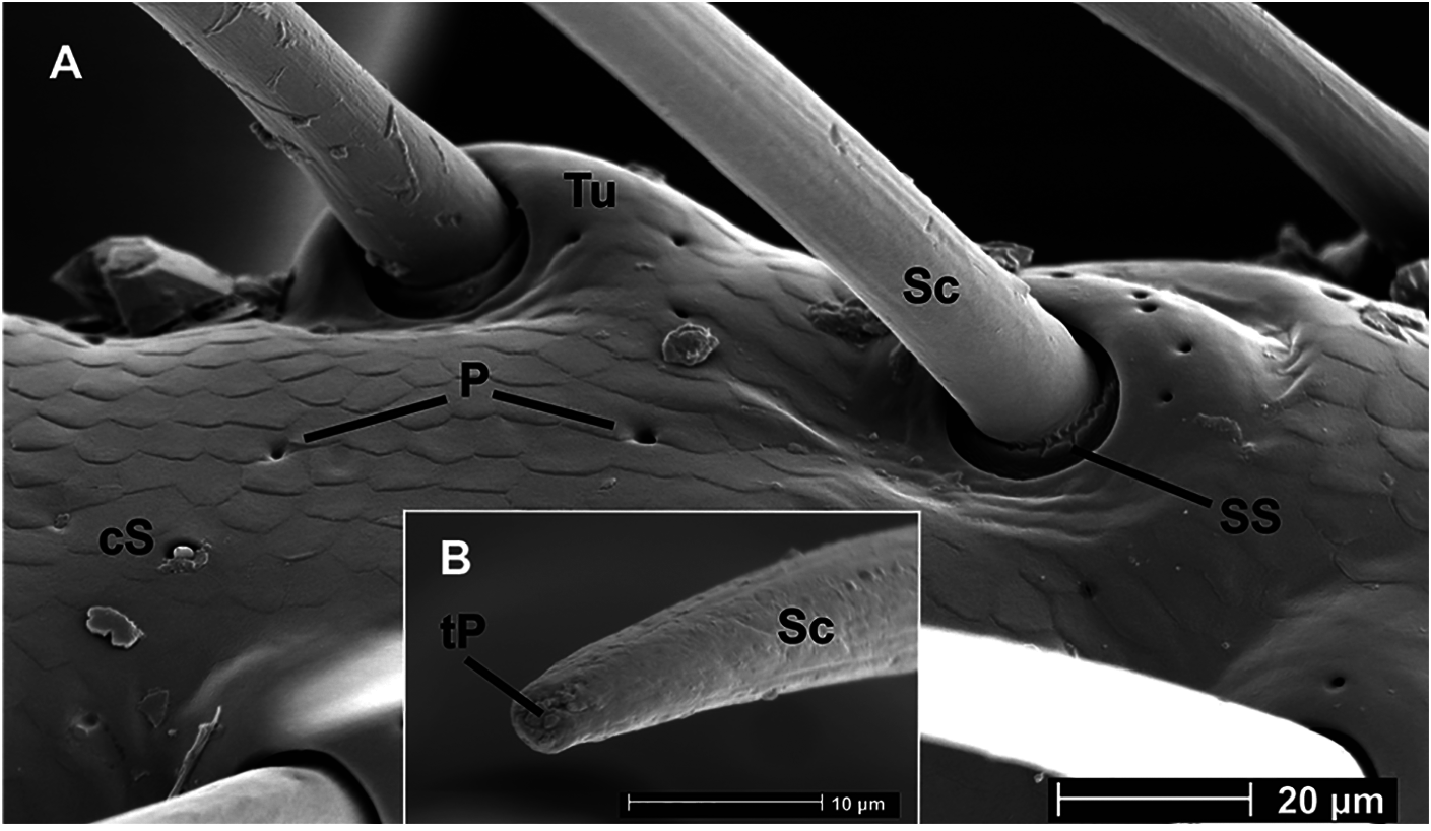
Under a scanning electron microscope SEM, the long setae of the berry-like stipes turned out to be truncate and open-pored ( Fig. 3 View Fig ) in contrast to the setae of a "normal" stipes, of which the tips are long and fine ( Fig. 4 View Fig ). The pore of the truncate seta tip may be filled with rests of a probable (dried) receptor lymph ( Fig. 3B View Fig ). Furthermore, the cuticle of the berry-like stipes is laced with (gland?) pores ( Fig. 3 View Fig ), whereas in the "normal" stipes the cuticle is free of pores ( Fig. 4 View Fig ); the function of the stipal pores is unknown. Campaniform sensilla as mechanoreceptors, which respond to stress and strain within the cuticle ( Gewecke 1995), are on both types of stipites.
The bow of truncate setae on the laterally bulging stipes in N. rubostipes sp. nov. may be part of a contact-chemosensitive system of food control, in contrast to N. impunctata sp. nov. with tapered setae on a "normal" stipes, where food control happens mechanosensitively. The pores on the cuticle of the stipes in N. rubostipes sp. nov. ( Fig. 3A View Fig ) point to a gland system, which is absent in N. impunctata sp. nov.
The lateral berry-like bulge of the stipes may enclose structures of a chemosensitive food control system and of a gland system in contrast to the flattened "normal" mechanosensitive stipes. The open-pored stipal setae ( Fig. 3B View Fig ) point to such a chemosensitive system. Contact-chemosensitive sensilla are well analyzed in spiders ( Drewes & Bernard 1976, Harris & Mill 1977, Vallet et al. 1998, Foelix 2015). They differ from common mechanosensitive setae by the open-pored tip, and they serve to test the chemical quality of substrates and thus the edibility of prey ("taste by touch"). Whereas in spiders the chemosensitive sensilla are located mainly on the forelegs and in a small number also on mouthparts, in Coleoptera chemosensitive setae on mouthparts are definitely plausible.
The analysis of the inner anatomy of the berry-like bulging stipes and of the supposed neurophysiological structures of a chemosensitive food control system and of a gland system was not the objective of the present paper. Our hypothesis may encourage to such an investigation. However, the berry-like bulge, hiding an evolutionarily complex system, is the only macroscopic character and is common to only a few related species within the genus Nebria .
We interpret this unique (and so far overlooked) berry-like stipes, observed in the two known Epinebriola species N. barbata and N. ganeshi (and? N. restricta ) as in three new species (described below), to be apomorphic, a character which is part of a so far unknown contact chemosensitive food control system within the genus Nebria . Due to the neurophysiological complexity which manifests phenologically in that conspicuous berry-like bulge, we state a serious evolutionary weightiness for this stipes character. We interpret this well defined species group to be monophylic, and therefore we propose the name Barbonebriola on subgeneric level for this species group. The fact of paraphyly in the subgenus Epinebriola allows to isolate the ganeshi species group without violating a possible monophyly in the remaining subgenus.
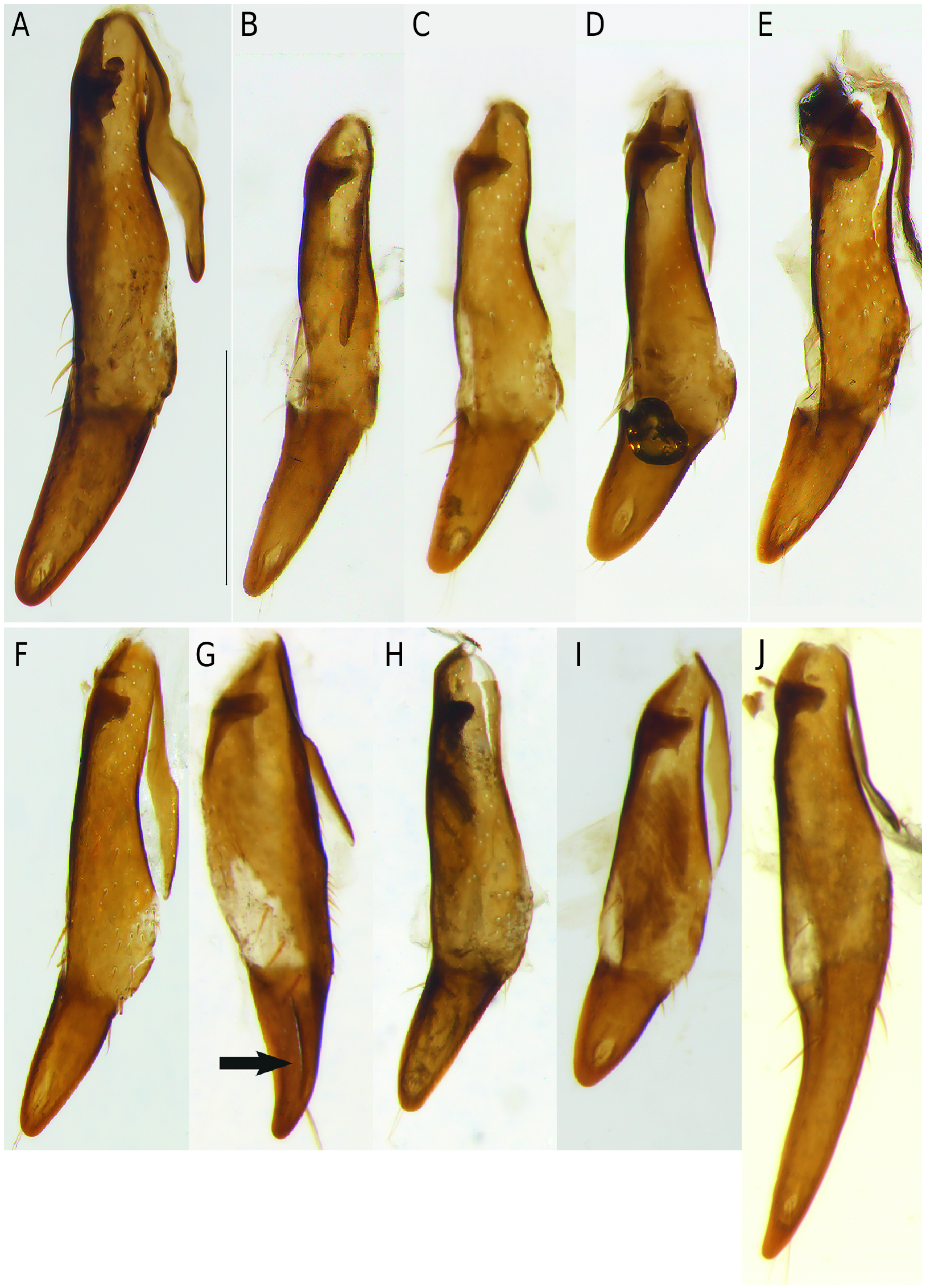
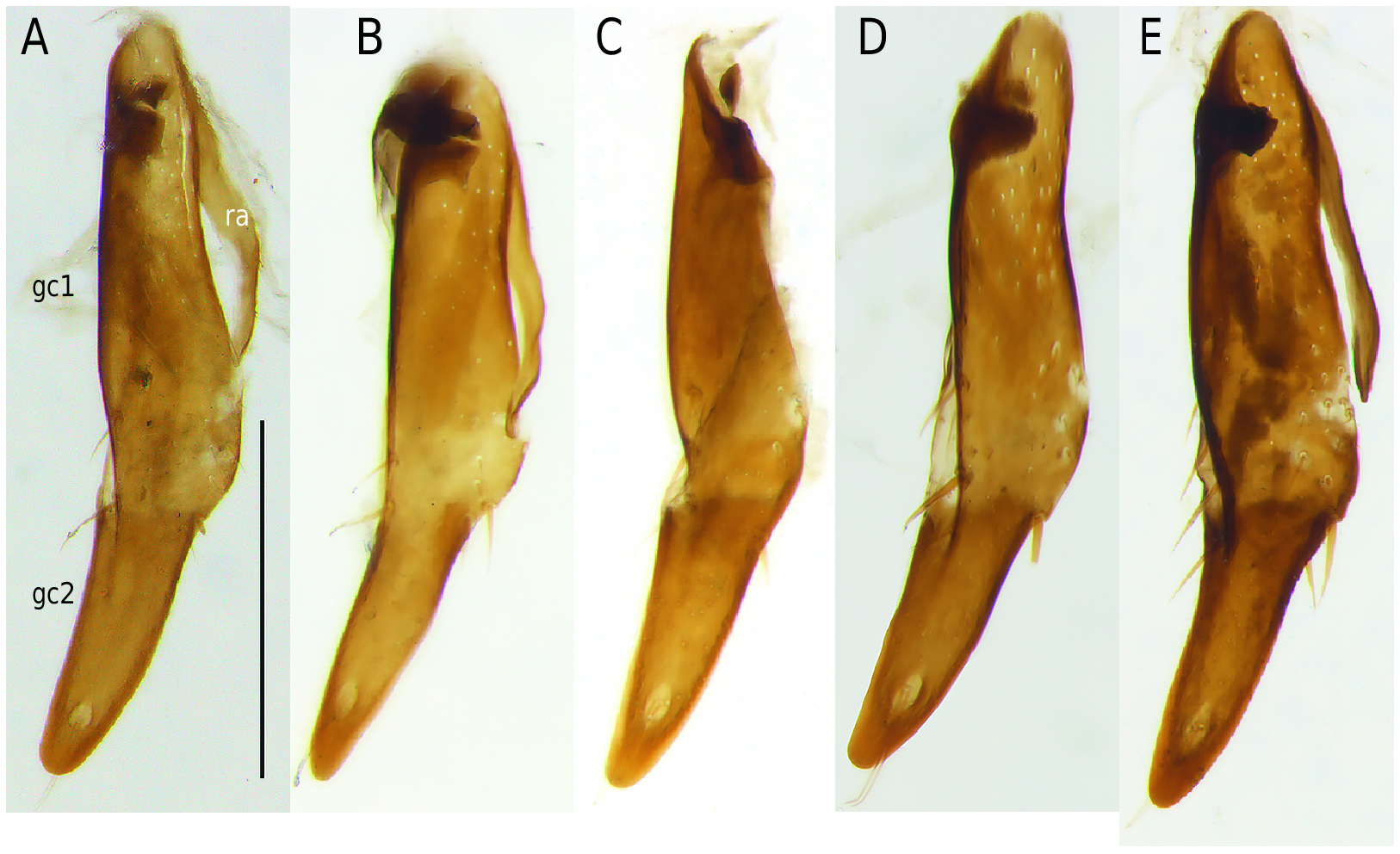
Gonocoxae (or coxostyli) of female genitalia of Nebria species are single or with two gonocoxites. The gonocoxae are rotated 90° in repose. In the subgenera Epinebriola and Barbonebriola sp. nov. the female gonocoxae are bipartite with a subcylindrical lightly sclerotized proximal gonocoxite 1 (or stylomere 1), and a highly sclerotized distal gonocoxite 2 (or gonostylus or stylomere 2). Gonocoxite 1 and 2 are continuously sclerotized ventrally giving the impression of unjointed or fused segments, whereas dorsally a membranous area points to a possible articulation between the gonocoxites ( Fig. 10G View Fig ). Up to now gonocoxae characters have been disregarded and unvalued in palearctic species of the genus Nebria . The present examination of the gonocoxae of female Epinebriola specimens shows an astonishingly wide variability in shape and length of the distal gonocoxite 2 (see Fig. 10 View Fig ), whereas in Barbonebriola subgen. nov. all species are characterized by a distinct uniformity in shape and length of the gonocoxite 2 ( Fig. 9 View Fig ). The shape of the gonocoxite 2 seems to be a stable character in the Barbonebriola species group, and its uniformity within Barbonebriola confirms the monophyly of this new subgenus.
Remark: Ledoux & Roux (2005) mentioned an unisetose apex of the gonocoxite 2 in most ("presque toujours") Nebria species. In the present analysis all examined gonocoxite 2 bear two closely adjoined preapical setae.
Fig. 6: Left anterior
angle of the pronotum. A: Nebria (Barbonebriola) barbata ANDREWES, 1929 : the anterior margination of the pronotum merges with apex of the anterior angle (arrow). B: Nebria (Epinebriola) montisanimae sp. nov.: the anterior margination of the pronotum merges with a ridge at the anterior angle (arrow); the ridge continues along the lateral groove to the lateral seta. Scale bar = 0.5 mm.
Barbonebriola subgen. nov. includes the species of the former ganeshi group sensu Ledoux & Roux (2005) and is defined by the following characters:
• Stipes of maxilla laterally berry-like bulging, with 6–10 tubercles bearing each a seta (not examined in N. restricta ).
• Penultimate labial palpomere trisetose.
• Head without lightened spots on the vertex.
• Anterior margination of the pronotum merged with the apex of the anterior angle ( Fig. 6A) (not examined in N. restricta ).
• Midlateral seta of the pronotum inserted in the lateral groove, never on the lateral edge, though disturbing the lateral outline of the pronotum. (Midlateral seta absent in N. tenuisulcata sp. nov.)
• Basolateral seta of the pronotum absent.
• Scutellar seta absent.
• Elytra with humeral carina.
• Intervals 3, 5 and 7 of the elytra asetose.
• Hindwings atrophied to a short strap-like vestige.
• 3 rd abdominal sternum with a row of 1–8 medial setae (not examined in N. restricta ).
• Edeagus of simple, coarse outline, less structured. Base wide. Mid-shaft thick, regulary rounded and progressively narrowed to the apex.
• Mid-shaft of the endophallus densely and extensively covered with long setae ( Fig. 8D View Fig ). This character was already reported by Ledoux (1984) for N. ganeshi . Not examined in N. restricta and N. tenuisulcata (only female holotype known).
• Gonocoxa: Gonocoxite 2 long, longer than of half length of gonocoxite 1, relatively slender at base, slightly narrowing near base, parallel or subparallel sided in median part, narrowing towards apex. Apex rounded, slightly arcuate dorsally, but dorsally not grooved. Gonocoxites ventrally unjointed and continuously sclerotized, dorsally separated by a membranous area. (Gonocoxa not examined in N. restricta ).
Diagnosis: The species of the subgenus Barbonebriola sp. nov. are recognizable by the laterally bulging and tuberculous stipes of the maxilla ( Fig. 4 View Fig , 5 View Fig ).
Etymology: The subgeneric epithet refers to the "hairy" outlook of the mouthparts (stipes and submentum) of all species of the new subgenus (barba, latin = beard). The gender of the genus-group name is feminine.
Remark: Andrewes (1929) did not etymologically explain the name of N. barbata . Due to the "hairy" stipes and its easy visibility in dorsal view, the name barbata may refer to this beard-like appearance of the head's side. Obviously the peculiar "hairy" stipes character was observed by Andrewes and even eponymously conserved in the species name barbata , but he did not make a corresponding note.
Monophyly: The monophyly of the Barbonebriola species group seems clear and is suggested by two characters: the consistent and exclusive bulging maxillary stipes (supposedly apomorphic), and the consistent shape of the distal gonocoxites 2 of the gonocoxae in females (see below).
Catalogue of species of Barbonebriola subgen. nov.:
Nebria (Barbonebriola) barbata ANDREWES, 1929 (transferred from subgenus Epinebriola )
Nebria (Barbonebriola) ganeshi LEDOUx, 1984 (transferred from subgenus Epinebriola )
Nebria (Barbonebriola) kagmara sp. nov.
? Nebria (Barbonebriola) restricta LEDOUx & ROUx, 2005 (transferred from subgenus Epinebriola ; uncertain assignment)
Nebria (Barbonebriola) rubostipes sp. nov.
Nebria (Barbonebriola) tenuisulcata sp. nov.
Distribution: The distributional area of the species of Barbonebriola subgen. nov. covers an area from Kashmir to Western Nepal Himalaya ( Fig. 14 View Fig ). The large distributional gaps in the Western Himalaya of India may be an artefact due to insufficient exploration and thus, other Barbonebriola species may be expected in this area.




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |