
Herentia thalassae David and Pouyet, 1978
|
publication ID |
https://doi.org/10.1080/00222930802109140 |
|
publication LSID |
lsid:zoobank.org:pub:2B6D5D3B-8F6D-4F0C-A377-784C4CBCF7E2 |
|
persistent identifier |
https://treatment.plazi.org/id/0395A723-FFBA-FFEA-FEE5-FDB5FF70EC27 |
|
treatment provided by |
Felipe |
|
scientific name |
Herentia thalassae David and Pouyet, 1978 |
| status |
|
Herentia thalassae David and Pouyet, 1978 View in CoL
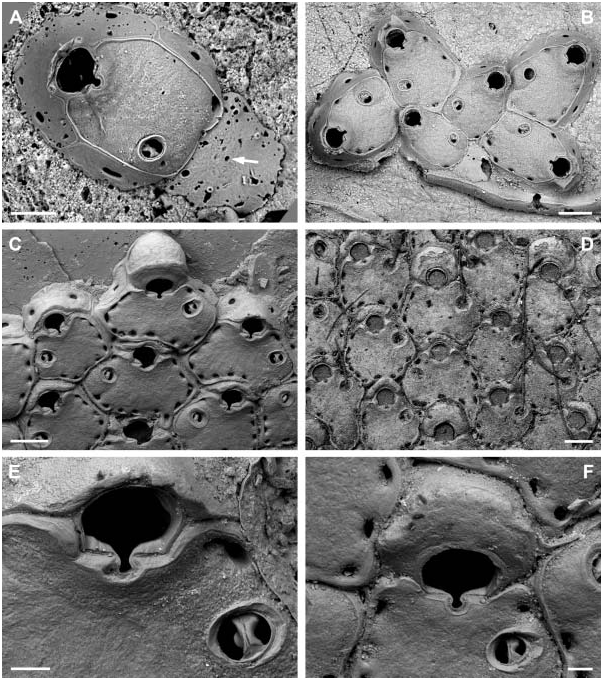
( Figure 4 View Figure 4 )
Herentia ( Herentia) thalassae thalassae David and Pouyet 1978 , part: p. 172, Plate 1, Figure 1 View Figure 1 ,?non 2.
Material examined
MNHN 7619 About MNHN , north-western Spanish shelf, ‘‘Thalassa’’ 1968, Stn 825 ( 42 ° 22,49N; 9 ° 24,89W), 520– 480 m, two colonies on rock GoogleMaps .
Measurements
ZL 828¡71, 730–941 (1, 14); ZW 698¡100, 565–906 (1, 14); OL 183¡10, 166–201 (1, 13); OW 156¡10, 133–170 (1, 13); ApL 179; ApW 174; OvL 352; OvW 457; AL 152¡12, 133–176 (1, 15); AW 125¡12, 101–138 (1, 15).
Description
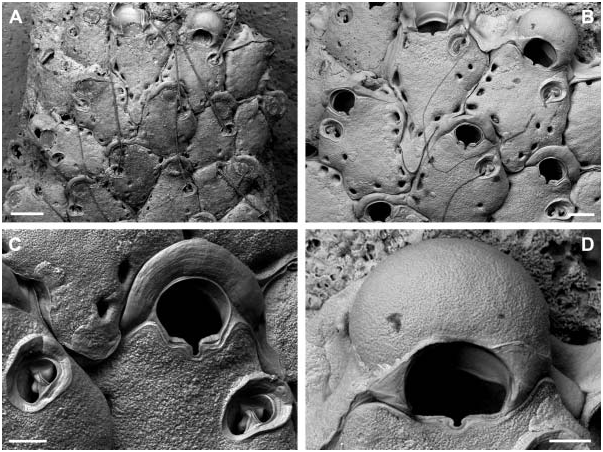
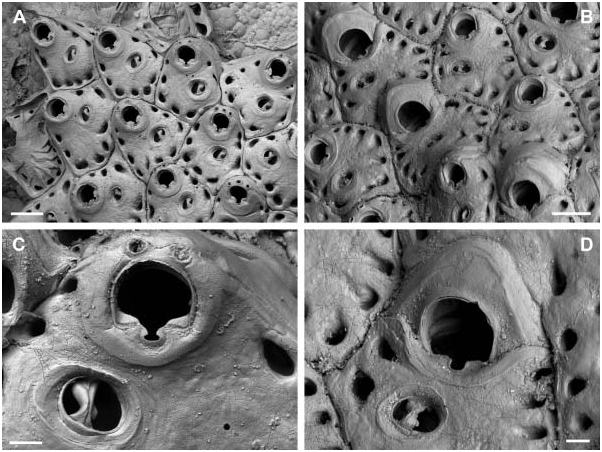
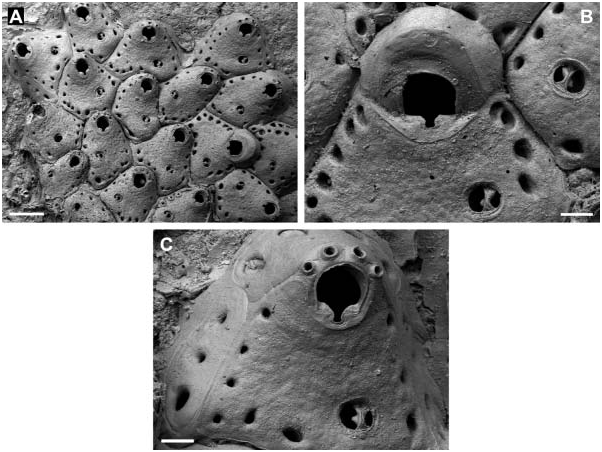
Colony encrusting unilaminar, initially multiserial, later oligoserial. Zooecia hexagonal to polygonal, usually distinctly broader proximally, separated by deep grooves; demarcated areas of vertical walls with two (in the two distal areas) and three to four (in lateral areas) round to oval communication pores of variable size. Frontal wall slightly convex proximally, distinctly rising distally towards an elevated orifice, surface rugose, imperforate except for a row of 8–15, round to oval, marginal pores of variable size. Orifice dimorphic, inner distolateral orifice rim with an immersed, very narrow shelf; primary orifice in autozooids slightly longer than broad and widest distal to mid-distance, anter horseshoe-shaped, proximal margin straight, sinus relatively broad and deep, ending in a transverse ellipse, condyles distinct, as long as proximal margin, narrowing towards sinus with an increasing slope-angle; aperture in ovicellate zooids larger, anter D-shaped, proximal margin wider with a slightly broader sinus, condyles distinctly narrower and almost straight. Primary orifice and aperture encircled by a smooth broad band of gymnocystal calcification. Three or four distal oral spines.
Ooecium initially spherical and tall, rising well above colony surface, later partly immersed by distal zooid; exposed surface smooth with a relatively narrow, curved, flattened rim at an acute angle to frontal plane, rim not covering the frontal area completely.
Avicularium single, situated proximal to mid-distance of zooecium (with two or three marginal pores between avicularium and orifice), at proximolateral zooecium margin or occasionally proximal to orifice at some distance from lateral wall, cystid continuous with frontal wall; rostrum oval with a broad, smooth and slightly raised rim sloping towards opesium, oriented distolaterally or laterally; mandible long and slender, setiform, up to twice the length of an autozooid; crossbar bilateral symmetrical, strong, slightly arcuate with the apex at centre, proximal margin straight or slightly concave when viewed from above except from a small subtriangular extension protruding from its basal centre, distal part of thick calcification set off from the proximal margin by a distinct line, calcification thinning medially, forming a small, U-shaped, distal opesium.
An ancestrula was not observed.
Remarks
The holotype of H. thalassae was presumably misplaced in the MNHN and could, unfortunately, not be found on request. We therefore re-described and figured another specimen from the same suite of the type location. From the short original description it is evident that H. thalassae is characterized by four oral spines which are present in the analysed specimen ( Figure 4C View Figure 4 ). However, it is questionable if the second specimen figured in the original description (David and Pouyet 1978, Plate 1, Figure 2 View Figure 2 ; collection number FSL 116270) belongs to the same species because in this SEM image there are no spine bases visible on the distolateral orifice margin and the areolar pores are considerably smaller.
Initially introduced as a subspecies to distinguish the recent taxon from the Pliocene H. thalassae iberica David and Pouyet, 1978 , H. thalassae differs sufficiently from the fossil to justify giving it full species rank. Judging from the poor photo given in David and Pouyet (1978, Plate 1, Figure 3 View Figure 3 ), the Mediterranean H. iberica is actually more closely related to H. hyndmanni and H. majae due to its rather distally positioned avicularium and should be compared with these taxa for synonymy. On the other hand, the presence of three or four spines in H. thalassae may suggest that it is a specimen of this species Hayward and Ryland (1979, Figure 82C; 1999, Figure 98C) figured as H. hyndmanni (see also Remarks there). While the spines were not preserved in the presently available material, Hayward and Ryland (1979, p. 196, 1999, p. 232) reported them to be very long and slender in their specimens.
Compared to H. hyndmanni , H. thalassae lacks the thickened rim framing the distolateral orifice margin, has three to four oral spines, and has a distinctly raised distal part of the zooid as well as a taller ovicell that projects significantly above the colony surface. It is furthermore distinguished by its more proximomedially positioned avicularium, and its ribbon-like colony growth, with each ribbon consisting of about four series of zooids spreading out in different directions from the multiserial colony centre. This growth type is, however, somewhat similar to the lobate growth type developed by Mediterranean deep-water colonies of H. hyndmanni , and may be an adaptation to deeper habitats (see Discussion).
Even in small colonies the proximal ancestral region was often iron oxide-stained and only the zooids near the active growth margin were pristine. This may suggest that either overall colony growth is relatively slow and that proximal zooids senesce or starve as new zooids are formed, and/or that the colonies are preyed upon. The latter assumption may be corroborated by the poor state of preservation and repair of some of the proximal zooecia in Figure 4A View Figure 4 (see Discussion).
H. thalassae View in CoL has so far only been recorded from relatively deep waters ( 480– 520 m) off northern Spain. As descriptions and illustrations are not given it is impossible to judge what Herentia species Jullien and Calvet (1903) and d’Hondt (1970, 1973, 1974, 1975) report from the eastern Atlantic without seeing the original material. Furthermore, some other specimens analysed here from the southern Portuguese outer shelf (MNHN 6764, 6760), co-occurring with H. hyndmanni View in CoL (see Remarks there), are distinctly smaller [ZL 597¡47, 524–683 (3, 9); ZW 442¡59, 352–560 (3, 9)], and have four to five oral spines in adult zooids, while the other morphological characters more or less correspond to the specimens from northern Spain. Yet, before a firm conclusion on the relationship between these specimens and H. thalassae View in CoL can be drawn, more material from southern Portugal needs to be analysed.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Herentia thalassae David and Pouyet, 1978
| Berning, Björn, Tilbrook, Kevin J. & Rosso, Antonietta 2008 |
H. thalassae
| David and Pouyet 1978 |
H. thalassae
| David and Pouyet 1978 |
H. hyndmanni
| : Norman 1909 |