
Parotocinclus halbothi, A & Lazzarotto & Reis, 2014
|
publication ID |
https://doi.org/ 10.1590/S1679-62252014000100002 |
|
DOI |
https://doi.org/10.5281/zenodo.4774110 |
|
persistent identifier |
https://treatment.plazi.org/id/0397C75C-FFBE-FFFB-C7C9-098E72F4389F |
|
treatment provided by |
Carolina |
|
scientific name |
Parotocinclus halbothi |
| status |
sp. nov. |
Parotocinclus halbothi View in CoL , new species
Holotype. MCP 48029, 19.4 mm SL, female, Brazil, Pará , Oriximiná , creek tributary to igarapé do Moura at Platô Monte Branco , rio Trombetas drainage, Amazon basin, 1º35’58.09”S 56º31’21.83”W, 21 Apr 2009, V. Reis & H. Lazzarotto. GoogleMaps
Paratypes. MCP 48030, 6 View Materials , 17.5-19.5 mm SL, collected with the holotype . MCP 48098, 1 View Materials c&s, 19.7 mm SL; and INPA 39890 View Materials , 4 View Materials , 18.5-19.9 mm SL + 1 immature unmeasured, same locality as holotype, 10 Oct 2009 GoogleMaps , V. Reis & M. Andrade . MPEG 17299 View Materials , 2 View Materials (juveniles), 11.4-15.5 mm SL, Brazil, Pará, igarapé 1500 on track 4, Estação Ecológica Grão-Pará, upper rio Mapuera, rio Trombetas basin, 1º16’20.8”N 58º41’09.2”W, 29 Aug 2008 GoogleMaps , F. R. Silva . USNM 409918 About USNM , 2 About USNM , 18.1-18.7 mm SL, Suriname, Sipaliwini, downstream waterfall in right tributary of upper Paloemeu River, Marowijne River basin, 2º27’21”N 55º37’35”W, 11 Mar 2012 GoogleMaps , J. Mol & K. Wan Tong You. USNM 408454 About USNM , 1 About USNM , 17.1 mm SL, Suriname, Sipaliwini, left tributary to upper Paloemeu River, 1 km downstream of basecamp, Marowijne River basin, 2º28’38”N, 55º38’17”W, 13 Mar 2012 GoogleMaps , J. Mol & K. Wan Tong You.
Diagnosis. Parotocinclus halbothi is distinguished from congeners of northeastern to southeastern Brazilian rivers and the río de La Plata basin in having the cheek canal plate elongated posteriorly on the ventral surface of head and contacting the cleithrum (vs. canal plate not elongated posteriorly and not contacting the pectoral girdle). It differs from species of Parotocinclus from the Guianas, Orinoco, and Amazon basins, except Parotocinclus collinsae , by the absence of a triangular patch of dark pigmentation on the anterior portion of dorsal-fin membrane (vs. the possession of such pigmentation), the possession of unicuspid accessory teeth on both the premaxilla and dentary (vs. accessory teeth absent), and by lacking the Y-shaped light mark on the snout and head (vs. possessing a Y-shaped light mark on the snout and head). The new species is distinguished from P. collinsae in having the small, circular, median abdominal plates arranged in 4 to 7 irregular longitudinal series (vs. median abdominal plates transversely elongated and arranged in one longitudinal series; Fig. 2 View Fig ), the snout and rostrum pigmentation in a dorsal view not reticulated and without dark dots on the dorsolateral surface of head (vs. snout and rostrum pigmentation reticulated and with dark dots present on head), and by the rudimentary adipose fin without a membrane (vs. the presence of an obvious adipose fin with a hyaline membrane).
Description. Proportional measurements in Table 1 View Table 1 . Dorsal profile of head from snout tip to parieto-supraoccipital tip convex, with small depression immediately posterior to snout tip, rostrum slightly depressed. Dorsal profile of body straight and posteroventraly slightly slanted from dorsal-fin origin to insertion of caudal fin. Trunk and caudal peduncle rounded to ovoid in cross section, slightly flattened ventrally and compressed caudally. Snout elongated, depressed, with its anterior margin rounded to slightly pointed in dorsal view. Body progressively narrowing posteriorly from cleithrum.
Head flat to slightly convex between orbits; dorsal margin of orbit slightly elevated. Eye positioned dorsolaterally, comparatively large with iris operculum present. Body entirely covered by dermal plates except for ventral surface of head around lips, area immediately surrounding pectoral- and pelvic-fin insertions, and area around anus. Dermal plates of body uniformly covered with recurved odontodes arranged in approximate longitudinal rows except in abdominal region, with odontodes irregularly distributed. Lateral plates arranged in five longitudinal series on trunk. Dorsal plate series complete, with 22-23 (22) plates; mid-dorsal series incomplete, with 8-9 (9) plates; middle series complete, with 21-22 (21) plates.Lateral line with anterior 7-8 (8) anterior plates bearing canal and followed by 1 (1) unperforated plate, then 11 (11) posterior plates bearing canal, and 1-2 (1) terminal plates without canal.Mid-ventral series incomplete with 14-15 (15) plates; series terminating between origin of anal and origin of adipose fin. Ventral series complete and continuous from pelvic-fin origin to caudal-fin base, with 16-17 (17) plates. Predorsal plates 3-4 (3). Preadipose azygous plates 1. Coracoid completely exposed ventrally. Cleithrum not exposed along anterior margin but contacting cheek plate; arrector fossa closed. Lateral abdominal plates 5-7; plates elongate, not clearly arranged in line between pectoral-fin axilla and pelvic-fin origin. Middle abdominal plates small, circular, and irregularly arranged in 4-7 longitudinal series covering most of abdominal surface between pectoral girdle and anal opening. Preanal shield not well developed, sometimes divided in 2 or 3 plates ( Fig. 2 View Fig ).
Posterior tip of parieto-supraoccipital with small patch of enlarged, slightly raised odontodes relative to those of remainder of head and predorsal area. Odontodes on head and trunk otherwise of uniform size and distribution, diverging on snout and converging to parieto-supraoccipital. Enlarged odontodes present on most of border of snout, especially on rostral and postrostral plates, and on infraorbital 1; enlarged odontodes curved and posteriorly oriented.
Dorsal-fin rays I,7. Dorsal-fin origin at vertical slightly posterior to anal-fin origin. Dorsal-fin spinelet present, plate-like and triangular with sharply pointed lateral margins. Dorsalfin locking mechanism non-functional. Adipose fin vestigial, without membrane attaching to body. Pectoral-fin rays I,6. Large spine slightly arched; tip of adpressed spine reaching between middle and distal third of pelvic fin. Pectoral-fin axillary slit present, with large opening ventral to tip of posterior process of cleithrum. Pelvic-fin rays i,5. Fin short, with tip of adpressed fin reaching to, or falling short of, anal-fin origin. Adult males with fleshy flap along posterodorsal margin of thickened first pelvic-fin ray. Anal-fin rays i,5. Caudal-fin rays i,14,i, upper and lower unbranched rays subequal.
Color in alcohol. Dorsal and lateral portions of head and trunk light orange-brown to pale yellow with ventral surface lighter. Head darker between orbits from nostrils to centrum of compound pterotic. Posterior portion of parieto-supraoccipital and compound pterotic conspicuously lighter than surrounding areas. Contra some congeners,Yshaped light mark absent on dorsal surface of snout and parieto-supraoccipital. Predorsal region slightly darker than background, but not forming transverse bar at anterior dorsalfin base and dark triangle at anterior dorsal-fin membrane absent. Trunk with two conspicuous brown bars, extending transversely from mid-dorsal region to lateral line. First bar situated between dorsal-fin base and adipose-fin origin; its anterior margin continuing as thinner bar two plates wide below lateral line, but falling short of ventral midline. Second bar on posterior portion of caudal peduncle and ending on caudal-fin base; both anterior and posterior margins of bar continue ventrally as thinner bars below lateral line to ventral midline. Fins with transverse, inconspicuous brown bands formed by concentration of chromatophores on rays; bands more visible on leading rays. Dorsal fin with 4-5 dark bands, first located at base of spine. Pectoral fin with 3-4, pelvic-, anal-, and adipose-fin spines with 1-2 dark spots. Caudal fin with 3 irregular, thin brown bands; third band best developed with posterior margin sometimes darker. Interradial membranes of all fins hyaline to light yellow.
Distribution. Parotocinclus halbothi is known from four localities, two streams in the rio Trombetas basin, a tributary to the rio Amazonas, and two in tributaries to the upper Marowijne River, in southern Suriname ( Fig. 3 View Fig ). The type locality is a small stream tributary to the igarapé do Moura, whose headwaters are located on the northern and western slopes of Monte Branco plateau, and drains into the lago Moura on the right bank of the rio Trombetas. Two other small (11.4 and 15.5 mm SL) juvenile specimens were collected inside the Estação Ecológica Grão-Pará, in a small stream tributary to upper rio Mapuera, rio Trombetas drainage, in the hills of the Brazilian-Guyanese border. Finally, two lots were recently collected in creeks tributary to the upper Paloemeu River, Marowijne River basin in Sipaliwini, southern Suriname, not far from headwater tributaries to the rio Trombetas and the Brazilian border.
Etymology. The specific epithet, halbothi , is given in memoriam to the Brazilian ichthyologist Dário Armin Halboth (29 June 1965 - 1 July 2003), an excellent field biologist and one of the first researchers to study the effects of bauxite tailings deposited in an Amazonian lake (lago Batata, a floodplain lake located in the floodplains of rio Trombetas, Pará State) on fish communities. Before his early death, Dário Halboth dedicated himself to describe the ecological features of the fishes living in streams of Amapá State, Brazil.
Remarks. Parotocinclus halbothi is possibly the sisterspecies to P. collinsae , as they share three characters not present in the remaining species of Parotocinclus in the Amazon-Guianas-Orinoco clade. These two species share the absence of a triangular patch of dark pigmentation on the anterior portion of the dorsal-fin membrane, the lack of a Y-shaped light mark on the snout and head, which are both present in all other species, and the possession of unicuspid accessory teeth on both the premaxilla and dentary, with such dentition absent in the remaining species. The phylogenetic value of these transformations, however, remains to be evaluated.
Parotocinclus halbothi is one of the smallest known hypoptopomatines, with the largest examined specimen being 19.9 mm SL. Despite being so small, examined specimens of P. halbothi are adults, as evidenced by being fully plated including complete abdominal plate covering ( Fig. 2 View Fig ) and by males having a fleshy flap along the posterodorsal margin of the thickened first pelvic-fin ray.
Ecological notes. The type locality of Parotocinclus halbothi ( Fig. 4 View Fig ) is a third order, clear water stream draining the northern slopes of the Monte Branco plateau into the lago Moura, a floodplain lake located on the right bank of rio Trombetas. The stream flows through a pristine, dense primary rainforest area, whose trees provide a 90% canopy cover over the stream, preventing sunlight from directly reaching ground level. No aquatic vegetation was observed at the type locality. The stream has an average width of 3.1 m and depths of up to 56 and 41 cm in the rainy and dry seasons, respectively. The water current in the mid-channel was fairly constant, with low-speed or still currents found only close to the stream margins or behind plant debris.Average water velocity ranged from 0.44 m /s to 0.76 m /s and water flow between 0.42 m 3 /s and 0.74 m 3 /s with both maximum values recorded during the rainy season. Some water features demonstrated slight variation between the rainy and the dry seasons: temperature (25.8-26.41 oC), pH (4.88-5.00), conductivity (10-11 µS/ cm 2) and Dissolved Organic Carbon (DOC, 3.02-3.11 mg /L). Dissolved Oxygen (DO) ranged from 6.16 mg /L to 7.6 mg /L, being higher in the dry season, and turbidity ranged between 3.1 NTU and 1.12 NTU, being higher in the rainy season. The substrate was composed primarily of fine white gravel (40-50%), coarse sand (20-40%), and leaf litter banks (18-30%). Individuals of P. halbothi were observed and collected in deeper parts of the main channel, mainly over the gravel substrate. Other fishes collected syntopically were: Aequidens pallidus , Apistogramma agassizi , A. eunotus , A. regani , Bryconops caudomaculatus , B. melanurus , Callichthys callichthys , Copella nigrofasciata , Erythrinus erythrinus , Gymnorhamphichthys rondoni , Helogenes marmoratus , Hoplias malabaricus , Mastiglanis asopos , Pyrrhulina aff. brevis , Rivulus dibaphus , and Synbranchus marmoratus .
The streams draining the plateaus of Saracá-Taquera National Forest in the Nhamundá and Trombetas region share a common set of features, such as clear waters, temperatures around 26 o C, low conductivity, pH around 5, high DO and low DOC content. Nonetheless, there is sufficient variation in structural features among streams that the composition of the local ichthyofauna can vary significantly ( Reis, 2011). For instance, among 45 sampling sites studied in 2009 in several microbasins, a fine gravel substrate such as that in which Parotocinclus halbothi was collected, was only found in the streams flowing north from the Monte Branco plateau. It is possible that the presence of this particular microhabitat is a crucial factor for the occurrence of P. halbothi in the lower Trombetas basin. Alternatively, the absence of P. halbothi in the other sampling sites may reflect the species’ historical distribution limits, since the majority of the studied streams flow south thereby draining into Nhamundá-Trombetas varzea lakes much further downstream than do the streams flowing north from the Monte Branco plateau.
We have no detailed ecological information on the collecting sites of the upper rio Mapuera or the Paloemeu River, except that in both cases the area was very well preserved and covered with original forest.
Conservation remarks. The type locality of Parotocinclus halbothi and adjacent streams with similar features originate in pristine primary rainforest areas on the slopes of the plateaus of Saracá-Taquera National Forest (a conservation unit) where officially permitted bauxite extraction takes place. The forest is totally removed from the top of the plateaus during the mining activity and replanted after the ore is removed. Whether that activity results in adverse impacts on the stream biota is currently under investigation. Nonetheless, the presence of the new species in a single stream with such specific habitat features in that area merits special attention in the conservation of the natural condition of the lower rio Trombetas.
Comparative material. Material examined in addition to that listed in Lehmann & Reis (2012): Parotocinclus amazonensis: MZUSP 10145, holotype. Parotocinclus bahiensis: MNRJ 31944, 1. Parotocinclus bidentatus: MNRJ 27962, holotype. Parotocinclus cristatus: MNRJ 32133, 1. Parotocinclus jimi: MZUSP 24576, 5. Parotocinclus jumbo: MZUSP 69513, holotype. MZUSP 69516, 1, paratype. Parotocinclus longirostris: MZUSP 88877, 2. Parotocinclus muriaensis: MNRJ 28528, holotype. Parotocinclus planicauda: MZUSP 69348, 1. Parotocinclus prata: MZUSP 68359, holotype. MZUSP 68360, 1, paratype.

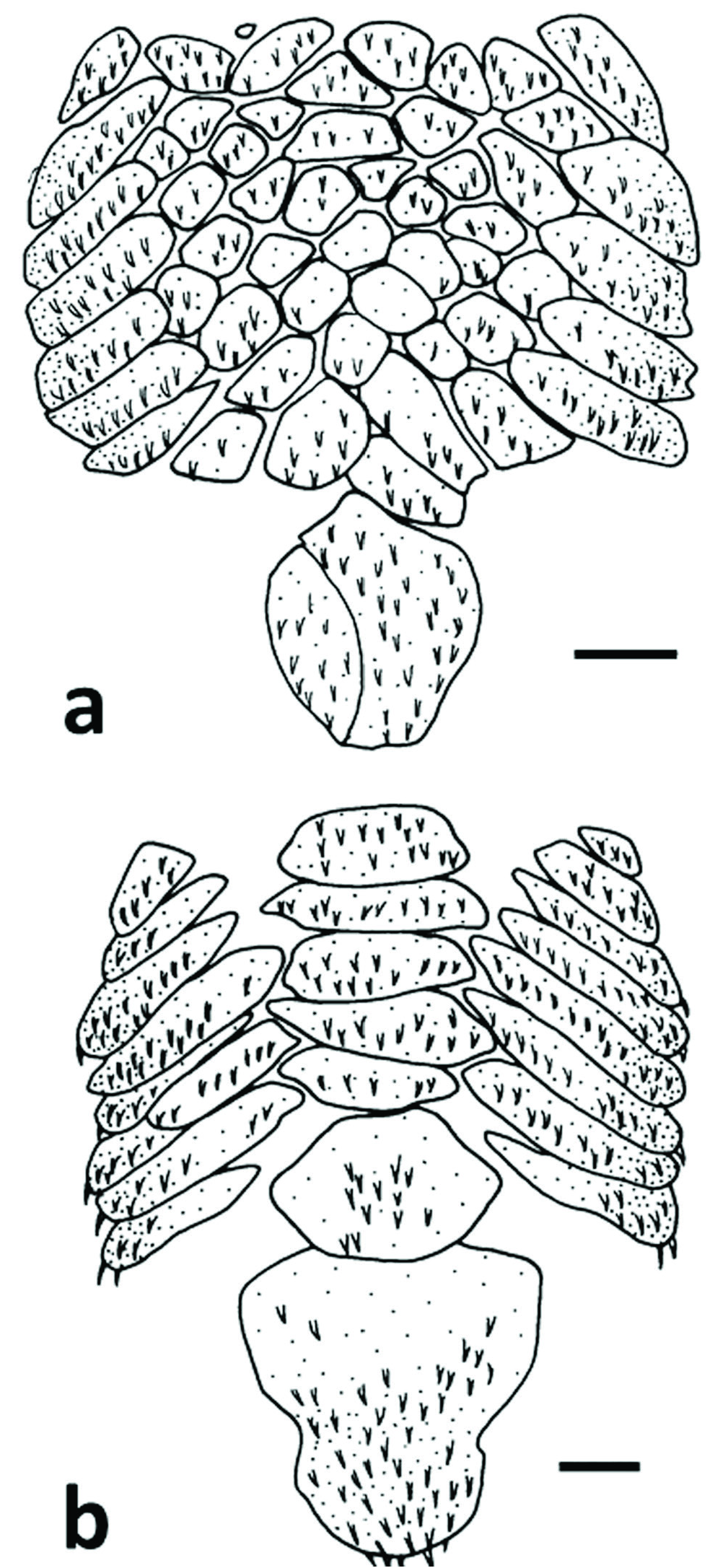


Table 1. Morphometrics of holotype (H) and paratypes (n = 12) of Parotocinclus halbothi as percents of standard length or head length. SD = standard deviation.
| Mesurements | H | Min | Max | Mean | SD |
|---|---|---|---|---|---|
| Standard length (mm) | 19.4 | 17.5 | 19.9 | 18.8 | - |
| Percent of Standard length | |||||
| Body depth | 15.9 | 14.7 | 19.5 | 16.3 | 1.1 |
| Predorsal distance | 51.3 | 51.0 | 55.5 | 53.0 | 1.4 |
| Prepelvic distance | 50.5 | 45.9 | 54.6 | 50.0 | 2.1 |
| Preanal distance | 70.4 | 66.2 | 75.2 | 69.6 | 2.4 |
| Preadipose distance | 79.7 | 78.1 | 86.0 | 80.7 | 2.3 |
| Dorsal-fin spine length | 19.9 | 17.8 | 21.6 | 20.0 | 1.1 |
| Anal-fin unbranched ray length | 13.2 | 13.0 | 17.3 | 14.9 | 1.2 |
| Adipose-fin spine length | 5.3 | 4.5 | 7.2 | 5.4 | 0.9 |
| Pectoral-fin spine length | 19.4 | 19.4 | 24.3 | 21.9 | 1.4 |
| Pectoral-fin spine depth | 1.3 | 1.3 | 2.0 | 1.7 | 0.2 |
| Caudal peduncle depth | 9.1 | 9.0 | 10.3 | 9.6 | 0.5 |
| Dorsal-adipose fin distance | 16.7 | 14.3 | 21.1 | 17.0 | 1.7 |
| Dorsal-fin base length | 15.1 | 12.1 | 15.1 | 13.1 | 0.8 |
| Lower caudal-fin principal ray | 23.7 | 21.9 | 26.5 | 24.2 | 1.3 |
| Cleithral width | 29.4 | 28.1 | 31.3 | 29.1 | 0.8 |
| Head length | 41.9 | 38.9 | 44.4 | 42.0 | 1.4 |
| Percent of Head length | |||||
| Head depth | 38.9 | 38.5 | 48.5 | 41.0 | 2.6 |
| Interorbital distance | 29.9 | 27.1 | 33.6 | 29.6 | 1.5 |
| Orbital horizontal diameter | 9.9 | 9.9 | 16.1 | 12.7 | 1.6 |
| Snout length | 58.0 | 57.8 | 65.6 | 60.0 | 2.5 |
| Internareal distance | 10.0 | 7.6 | 11.0 | 9.0 | 1.0 |
| MCP |
Pontificia Universidade Catolica do Rio Grande do Sul |
| V |
Royal British Columbia Museum - Herbarium |
| R |
Departamento de Geologia, Universidad de Chile |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |