
Liolaemus tregenzai, Pincheira-Donoso, Daniel & Scolaro, Alejandro, 2007
|
publication ID |
https://doi.org/10.5281/zenodo.176298 |
|
DOI |
https://doi.org/10.5281/zenodo.6237641 |
|
persistent identifier |
https://treatment.plazi.org/id/0399D46D-FFA7-3B63-FF36-80DC7A108656 |
|
treatment provided by |
Plazi |
|
scientific name |
Liolaemus tregenzai |
| status |
sp. nov. |
Liolaemus tregenzai sp. nov.
Figs. 1–2 View FIGURE 1 View FIGURE 2
Type Material. Holotype. MLP.R. 5274, adult male. Collected in Copahue Volcano slopes (37°50’S; 71°06’W), 2150 m, near Termas del Copahue, Departamento de Ñorquín, Neuquén Province, Argentina, by J.A. Scolaro, N. Scolaro, G. Scolaro and F. Tavera, February 6–10, 2006.
Allotype. MLP.R. 5275, adult female. The same data as holotype.
Paratypes. MLP.R. 5276-5277, both adult females. The same data detailed for the holotype and allotype. CHDPD 0 1098, adult male, CHDPD 0 1099, adult female. The same data detailed for the holotype and allotype. JAS-DC 806, adult male, JAS-DC 807, adult female. The same data detailed for the holotype and allotype.
Etymology. This new lizard species is named Liolaemus tregenzai after Tom Tregenza (Royal Society Research Fellow, University of Exeter), in recognition of his contributions to evolutionary biology. This dedication is also a personal recognition for the time spent guiding DP-D through a doctoral period of research in the United Kingdom.
Diagnosis. A large Liolaemus species belonging to the chiliensis clade diagnosed by Etheridge (1995), because it has a low number (4-4) of narrow supralabial scales, the last one elongated and upturned in the posterior margin, without a bladelike process on the posterior distal tibia, without femoral patch of enlarged scales, without tridentate dorsal scales, and with Meckel’s groove fused (see also Laurent 1992; Schulte et al. 2000; Lobo 2001; Pincheira-Donoso & Núñez 2005). All these traits differentiate L. tregenzai from L. periglacialis , L. kolengh , L. lineomaculatus and L. silvanae , which exhibit the diagnostic traits detailed for the lineomaculatus clade (see Donoso-Barros & Cei 1971; Cei 1986; Etheridge 1995). Etheridge (1995) recognized four or fewer precloacal glands in males of most of the species belonging to the chiliensis clade. In Liolaemus tregenzai these glands are entirely absents, which differentiate this species from almost every taxa into this clade, except from L. coeruleus , L. cristiani , L. flavipiceus , L. neuquensis , and L. thermarum , which do not exhibit precloacal glands. Liolaemus tregenzai differs from L. cristiani in having a Patagonian distribution, and green-bluish ventral coloration with intense black pigment on the throat, chest and abdomen, whereas L. cristiani occurs in the Andes of central Chile, and exhibit a brown-yellowish or copper-yellowish ventral coloration, with total absence of intense black pigmentation in both sexes. Also, in L. tregenzai sexual dichromatism is evident, being absent in L. cristiani . Significant differences in the colour pattern differentiate L. tregenzai from L. flavipiceus . The olive, brown-blackish or chest-nut dorsal coloration and the green-bluish ventral colour with intense black pigmentation characteristics of L. tregenzai , contrast with the yellow dorsal background, black on the head, flanks, throat and chest, and with the greyish or reddish ventral colour with scattered orange or reddish spots observed in L. flavipiceus . Liolaemus tregenzai also differs from L. thermarum , because this last species does not exhibit black pigmentation on the ventral surface, which is whitish with conspicuous pinkish shades. The Liolaemus species lacking precloacal glands that exhibit more phenotypic and geographical similitude with L. tregenzai are L. coeruleus , from Zapala and Pino Hachado ( Cei 1986; Pincheira-Donoso 2003) and L. neuquensis , from Copahue ( Cei 1986; Videla & Cei 1996), the same area where the new species occurs (see above). Liolaemus tregenzai differs from L. coeruleus in having a body size ranging between 63.4–90.2 mm (see below for details) which contrasts significantly (see above) with the 44.7–60.2 mm in L. coeruleus . Also, the number of scales around the midbody (and therefore, the proportional size of body scales in relation to body size) is significantly different between these species, having L. tregenzai a mean value of 80.63, whereas the mean value in L. coeruleus is 63.4. In addition, while L. tregenzai exhibits a green-bluish ventral coloration, the abdominal and pectoral background in L. coeruleus is blue or sky-blue. Finally, L. tregenzai differs from L. neuquensis in having a significantly larger body size (55.3–61.1 mm in L. neuquensis ; see above), and because L. neuquensis shows a prominent hemigular fold, completely absent in L. tregenzai . The same ventral differences detailed for L. coeruleus in relation to L. tregenzai , are valid for L. neuquensis .
Description of the Holotype ( Fig. 1 View FIGURE 1 ). Adult male, snout-vent length 82.5 mm, forelimb length 32.3 mm, hind limb length 47.9 mm. Tail broken. Head length 18.0 mm, head width 16.5 mm, head height 9.2 mm. Stout body, with prominent head. Hind limb adpressed reaches the shoulder. Neck wider than head. Lateral cervical folds, postauricaular, supra-auricular, longitudinal neck, oblique neck, and antehumeral, prominent, forming a V behind the external auditory meatus. Deep prehumeral pocket. Lateral nuchal scales small, granular, convex, nonoverlapping, with numerous tiny granules in the interstitial spaces. Hemigular fold entirely absent.
Upper head scales smooth, pitted with numerous sensitive organs mainly on snout. Rostral shield pentagonal, about four times as wide as high, and as wide as mental, directly surrounded by six scales (two supralabials, two lorilabials and two postrostrals). Two postrostrals, together with anterior supralabials almost separate nasals from rostral. Nasal shields longer than wide, as large as postrostrals, surrounded by six scales.
Nostril rounded, slightly over one half length of nasal, posterior in position. Four internasal scales, the medial pair as large as postrostrals. Two irregularly shaped azygous forming a longitudinal row on the snout, the anterior twice as large. The posterior in direct contact with a prefrontal azygous. A frontal azygous, slightly larger than interparietal. Two postfrontal shields, smaller than interparietal. Interparietal shield small, as large as nasals, pentagonal, slightly enlarged, flanked posterolaterally by a pair of irregularly shaped parietal scales relatively larger than frontal azygous, and anteriorly in direct contact with postfrontals. Parietal eye small and conspicuous, posterior in position. Supratemporal scales relatively convexes, smooth, irregularly shaped and exhibiting high variation in size. Temporal scales more regulars, weakly keeled and subimbricate or juxtaposed. Circum orbitalis conspicuous, formed by 13-13 small and often hexagonal scales. Supraoculars 6-4, strongly hexagonals and transversally expanded, up to eight times larger than circum orbitalis scales. Three rows of smaller scales disposed between supraorbitals and superciliars. Superciliar scales longitudinally expanded, imbricate. Canthal margin inconspicuous. Loreal region weakly deep, covered by 5-5 irregularly shaped scales which exhibit high variability in size. Ciliar scales do not form palpebral “combs”. Upper ciliar scales 15-15, lower ciliars 11-10. A single and longitudinally enlarged subocular shield. Supralabial scales 4- 4, between two and three times longer than high, the fourth scale is evidently upturned on the posterior margin. Between subocular and supralabials there is a single row of lorilabial scales whose size is similar or slightly smaller than supralabials. External auditory meatus as large as the eye surface, oval, and higher than wide. Tympanic scales 3-3, convex, slightly projected backwards over external auditory meatus. Auricular scales 2-2, weakly convexes, as large as tympanics.
Mental shield about twice as wide as high, surrounded by the anterior infralabials and the anterior postmentals, but not in contact with anterior sublabials. Two divergent and decreasing rows of 5-5 postmental scales. Infralabial shields 5-5, between two and three times wider than high, whose surface is similar or slightly larger than supralabials. A single row of sublabial scales between the anterior infralabials and the anterior postmentals. Gular scales smooth, flat, imbricate, with rounded margin.
Dorsal body scales small, whose surface represents 45–60% of the interparietal shield, subtriangular, subimbricate, with a conspicuous but blunt keel, and without mucrons. There are scarce tiny and granular scales in the interstitial spaces. Dorsal scales grade laterally into slightly smaller scales, being fundamentally similar, but with abundant tiny granules in the interstices. Scales surrounding forelimb insertion small, convex, granular and juxtaposed. Ventral body scales 1.5–2 larger than dorsal scales, rounded, smooth, flat, and imbricate. Scales around midbody 81.
Forelimb scales as large as the dorsal body scales, subtriangular, imbricate and slightly keeled, becoming smooth on the dorsal surface of the hand. Lower humeral region covered by small, convex, granular, and juxtaposed, becoming larger, rounded, subimbricate and weakly keeled on the forearm. Several scales on this zone exhibit a tridentate margin. Infracarpal scales smaller, strongly imbricate, with well developed keels, and often with a tridentate margin. Subdigital lamellae laterally expanded, with three prominent keels, often with three conspicuous mucrons in the margin, although several of them can exhibit up to five mucrons. Posterior surface of thighs with tiny, granular, convex and juxtaposed scales. Upper hind limb scales slightly larger than dorsal scales, subtriangular, subimbricate, and weakly keeled on the femoral area, grading into more rounded, more keeled and subimbricate-juxtaposed on the tibial area. Supratarsal scales almost smooth, triangular and imbricate. On the lower femoral surface (infrafemorals), scales are smooth, flat, rounded and imbricate, grading into slightly larger scales on the tibial zone (infratibials). Infratarsal scales small, strongly imbricate, with well developed keels, and with a tridentate margin. Subdigital lamellae laterally expanded, with 3–4 strong keels, and with up to six prominent mucrons on the margin. Subdigital lamellae on the fourth toe of the left foot 26 (non-measurable on the right foot). Upper caudals larger than dorsal body scales, keeled and imbricate. On the ventral surface, scales are triangular, slightly keeled and imbricate. Without precloacal glands.
Coloration. Based in the whole type series, including males and females. A sexually dichromatic species ( Figs. 1 View FIGURE 1 and 2 View FIGURE 2 ). Although the coloration pattern is fundamentally similar in both sexes, pigments in males are more conspicuous.
Males. Background coloration of dorsum and upper head surface dark green olive, with disperse light green scales. Some specimens can exhibit a brown-blackish general colour on the upper head scales. On the back sides the ground colour becomes darker, forming a kind of wide lateral band, which can be brown-blackish or almost completely black in some individuals. The lateral fusion of some small blackish spots can form transversal and fine stripes which contact the dark band of the flanks. Upper surface of limbs ranging from green olive to dark brown, with abundant, contrasting and small light green spots irregularly dispersed. Lower surface of limbs black. Dorsal surface of tail with dark green semilunar spots surrounded by inconspicuous light green shades, black on the lower surface. Background colour of ventral surface light and shiny greenblueish. This coloration reaches the lower flanks. On the throat, chest, and medial surface of abdomen completely and intensely black.
Females. Dorsal head surface dark green, with a few small light green spots. Dorsal background colour dark chest-nut, with abundant and irregularly dispersed light green scales and irregularly shaped dark brown or blackish spots. On the anterior back sides there is a relatively wide dark brown or blackish lateral band. Background colour of ventral surface pale green-bluish, with abundant and irregularly dispersed light brownish or brown-copper, sometimes forming incipient transversal stripes. The medial colorations of throat, chest and abdomen are dark grey or greyish, never black as in males. Upper surface of limbs and tail colour dark chest-nut with abundant light green and blackish small spots. In juveniles the coloration is more similar to females, with dark greyish ventral surface.
Variation. Based on the entire type series. Variation in allometric proportions revealed the existence of sexual dimorphism in most of the studied variables. Snout-vent length differs significantly between both sexes (t -test, t 9 = 2.978, P = 0.016), being 75.7–90.2 mm (mean = 82.6 mm; SD = 5.94) in males, and 63.4–80.8 mm (mean = 71.41 mm, SD = 6.02) in females. Head length shows also significant sexual differences (t -test, t 9 = 2.884, P = 0.018), ranging between 17.3–20.5 mm (mean = 18.83 mm, SD = 1.45) in males, and between 14.5–18.9 mm (mean = 16.29 mm, SD = 1.38) in females. Head weight differs significantly between sexes (t - test, t 9 = 2.828, P = 0.02), being 13.1–16.5 mm (mean = 15.2 mm, SD = 1.6) in males, and 12.4–14.4 mm (mean = 13.2 mm, SD = 0.79) in females. Sexes differ in head height (t -test, t 9 = 2.414, P = 0.039), ranging between 9.1–12.2 mm (mean = 10.2 mm, SD = 1.44) in males, and between 7.1–9.3 mm (mean = 8.51 mm, SD = 0.91) in females. Conversely, tail and limb lengths do not show sexual dimorphism. Tail length in males is 8.46 mm (only one specimen with the entire tail), and 93.5–105.4 mm (mean = 95.1 mm, SD = 1.69) in females. Forelimb length in males is 27.8–32.3 mm (mean = 29.38 mm, SD = 2.05) in males, and 23.8–30.8 mm (mean = 26.62 mm, SD = 2.8) in females (P = 0.123). Hind limb length is 41.8–47.9 mm (mean = 44.58 mm; SD = 2.73) in males, and 35.9–46.0 mm (mean = 40.56 mm, SD = 3.35) in females (P = 0.073). When contrasting the derived variables forelimb length/snout-vent length (t -test, P = 0.351; 0.36 ± 0.03 in males, versus 0.37 ± 0.02 in females) and hind limb length/snout-vent length (t -test, P = 0.256; 0.54 ± 0.04 in males, versus 0.57 ± 0.03 in females) they do not show significant differences between sexes.
Variation in scale characteristics shown indistinctly for both males and females. Number of supraocular scales 5–6 (mean = 5.33, SD = 0.52). Scales forming circum orbitalis 13–14 (mean = 13.33, SD = 0.51). Upper ciliar scales 13–15 (mean = 13.83, SD = 0.75). Lower ciliar scales 10–14 (mean = 11.67, SD = 1.63). Number of loreal scales 5–9 (mean = 6.71, SD = 1.38). Number of supralabial scales 4– 4 in the whole sample. Number of tympanic scales 2–4 (mean = 3.0, SD = 0.63). Number of auricular scales 2–3 (mean = 2.33, SD = 0.51). Number of postmental scales 4–6 (mean = 4.83, SD = 0.75). Number of infralabial scales 5–7 (mean = 6.0, SD = 0.63). Subdigital lamellae on the fourth toe of the left foot 24–26 (mean = 24.75, SD = 0.96; we counted lamellae on the left food because in the holotype the right one is damaged). Variation in dorsal body scales is observed in form and disposition, ranging from rhomboidal to subtriangular ( Fig. 3 View FIGURE 3 a, b), being juxtaposed-subimbricate to subimbricate-imbricate. Ventral body scales do not exhibit remarkable variation in relation to the holotype ( Fig. 3 View FIGURE 3 c, d). Number of scales around the midbody 71–85 (mean = 80.63, SD = 4.27). Precloacal glands are absent in males and females of the whole sample ( Fig. 3 View FIGURE 3 e, f).
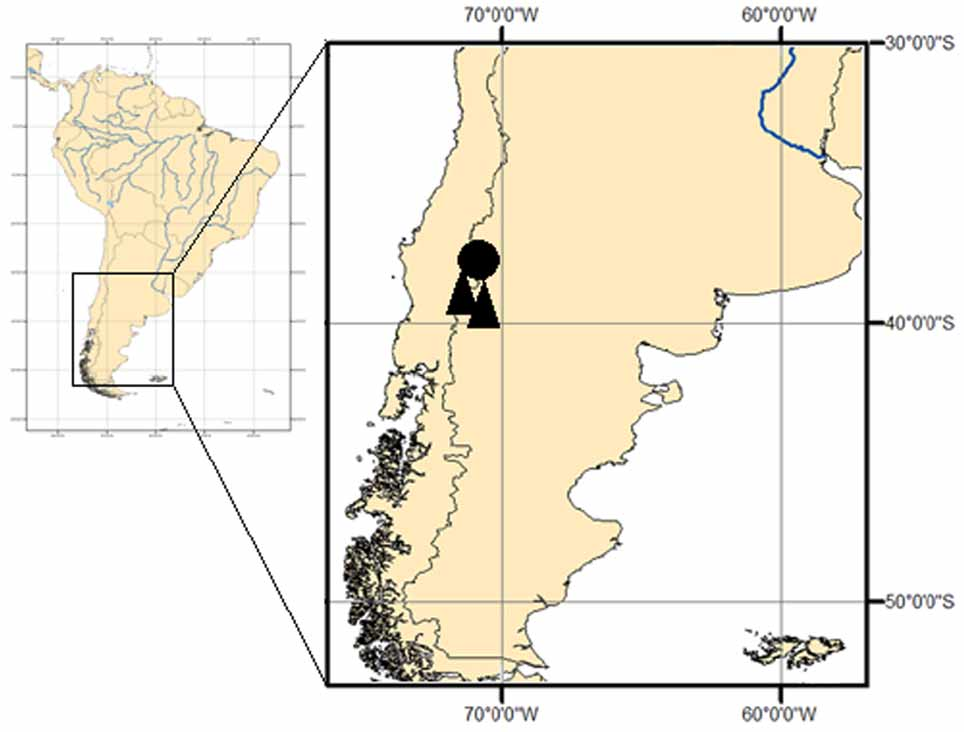
Distribution. Liolaemus tregenzai is known from Copahue Volcano slopes (37°50’S; 71°06’W), 2150 m, near Termas del Copahue, Departamento de Ñorquín, Neuquén Province, in north-western Patagonia of Argentina ( Fig. 4 View FIGURE 4 ).
Natural History. Liolaemus tregenzai is a Patagonian and diurnal species. Occurs in the Andean ecotonal zones between open antarctandic forests formed by Araucaria ( Araucaria araucana ) and areas recovered by Chasquea sp. It is a common species in areas characterized by clay sediments on the slopes of the Copahue volcano. Remarkably, L. tregenzai has often been found in the neighbourhoods of snow deposits, and close to thawing snow, which frequently form streams.
The diet of L. tregenzai is almost completely herbivorous. One of the studied specimens kept in captivity during more than nine months, was exclusively fed with lettuce. Nothing is known about other aspects of its natural history, remaining a mystery its reproductive mode.



| MLP |
Museo de La Plata |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.