
Ateleopus japonicus Bleeker 1853
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4027.3.4 |
|
publication LSID |
lsid:zoobank.org:pub:233FCD0C-1E48-498A-94C0-D00CD17322A9 |
|
DOI |
https://doi.org/10.5281/zenodo.6121936 |
|
persistent identifier |
https://treatment.plazi.org/id/039B9515-FFB6-FFCC-11ED-F88BFCD9418C |
|
treatment provided by |
Plazi |
|
scientific name |
Ateleopus japonicus Bleeker 1853 |
| status |
|
Ateleopus japonicus Bleeker 1853 View in CoL
[Japanese name: Shachiburi] ( Figs. 1–9 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8 View FIGURE 9 ; Tables 1–2)
Ateleopus japonicus Bleeker 1853: 19 View in CoL (based on the description of Temminck & Schlegel 1846); Temminck & Schlegel 1846: 255, pl.112, fig. 1 (description, type locality: Entrance of the large bay in the Oomura district, Nagasaki [prefecture], Japan [= Sasebo Bay]); Sauter 1905: 234 (key to species); Howell Rivero 1935: 93 (description, Japan); Okada & Matsubara 1938: 62, pl. 9, fig. 3 (key to species, Japan); Matsubara 1955: 272, pl. 25, fig. 91 (key to species, Japan); Shen & Ting 1972: 24, fig. 5 (description, Taiwan); Mochizuki 1984: 115, pl. 100 (description, Japan); Kulbicki et al. 1994: 14 (checklist, Chesterfield Islands, midway between Australia and New Caledonia); Randall & Lim 2000: 591 (checklist, South China Sea); Mochizuki 2001: 109 (description, Japan); Shinohara et al. 2001: 298 (checklist, Kochi, Japan); Aizawa 2002: 347 (key to species, Japan); Amaoka & Kobayashi 2003: 109, figs. 1–3 (description of postlarva, Yamaguchi, Japan); Shinohara et al. 2005: 407 (checklist, Ryukyu Islands, Japan); Bray & Hoese 2006: 457 (checklist, Australia); Shinohara et al. 2009: 700 (checklist, Tohoku district, Japan); Fricke et al. 2011: 358 (checklist, New Caledonia); Aizawa & Doiuchi 2013: 410 (key to species, Japan); Okiyama 2014: 269 (description of larva, Japan).
Ateleopus schlegelii van der Hoeven 1855: 326 (based on the description of Temminck & Schlegel 1846, type locality: Entrance of the large bay in the Oomura district, Nagasaki [prefecture], Japan [= Sasebo Bay]).
Ateleopus purpureus Tanaka 1915: 565 View in CoL (type locality: Ibaraki, Japan); Tanaka 1916: 396, pl. 107, fig. 328 (description, Ibaraki, Japan); Howell Rivero 1935: 96 (description, Japan); Okada & Matsubara 1938: 62, pl. 9, (key to species, Japan); Matsubara 1955: 273 (key to species, Japan); Okamura 1985: 439, fig. 218 (description, southern Japan, East China Sea); Xu 1988: 145, fig. 115 (description, East China Sea); Shinohara et al. 2001: 299 (checklist, Kochi, Japan); Aizawa 2002: 347 (key to species, Japan); Shinohara et al. 2005: 407 (checklist, Ryukyu Islands, Japan); Prokofiev 2006: 342 (description, Japan); Shinohara et al. 2009: 701 (checklist, Tohoku district, Japan); Aizawa & Doiuchi 2013: 410 (key to species, Japan).
Ateleopus tanabensis Tanaka 1918: 223 View in CoL (type locality: Wakayama, Japan); Howell Rivero 1935: 96 (description, Japan); Okada & Matsubara 1938: 62 (key to species, Japan); Matsubara 1955: 273 (key to species, Japan); Randall & Lim 2000: 591 (checklist, South China Sea); Mochizuki 2001: 109 (description, Japan); Shinohara et al. 2001: 299 (checklist, Kochi, Japan); Aizawa 2002: 347 (key to species, Japan); Kitagawa et al. 2008: 28 (description, Japan); Shinohara et al. 2009: 701 (checklist, Tohoku district, Japan); Aizawa & Doiuchi 2013: 410 (key to species, Japan).
Lectotype. RMNH 1410 (800 mm SL; Fig. 2 View FIGURE 2 ), Entrance of the large bay in the Oomura district, Nagasaki (prefecture), Japan (= Sasebo Bay).
Other materials. 77 specimens (212–820 mm SL): ZUMT 8320 (holotype of Ateleopus tanabensis Tanaka 1918 , 345 mm SL; Fig. 1 View FIGURE 1 ), Tanabe, Wakayama, Japan; ZUMT 8321–8323 (3 paratypes of A. tanabensis , 212–243 mm SL; Fig. 1 View FIGURE 1 ), Tanabe, Wakayama, Japan; CAS 224559 (2, 363– 456 mm SL), Tashi, Yilan, northeastern Taiwan, bottom trawler; CAS 235639 (298 mm SL), between Luzon island and Mindoro island, Philippines; FKUN 11585, 21863–21865, 21867, 22639–22640 (7, 325– 638 mm SL), Owase, Mie, Japan, bottom trawler; FKUN 32626 (227 mm SL), Kiinagashima, Mie, Japan, bottom trawler; FKUN 33359–33363 (5, 316– 565 mm SL), Suruga Bay, Shizuoka, Japan; HUMZ 35540–35541 (2, 423– 427 mm SL), off Cape Ashizuri, Kochi, Japan; HUMZ 49542 (440 mm SL), Tosa, Kochi, Japan, 180 m, bottom trawler Kousei Maru; HUMZ 49815, 105809, 109326 (3, 413– 427 mm SL), Mimase Fishmarket, Kochi, Japan; HUMZ 52245 (543 mm SL), Miya Fishmarket, Gamagoori, Aichi, Japan; HUMZ 72942, 106481 (2, 723– 764 mm SL), Fish Festival in Odawara, Kanagawa, Japan; HUMZ 106766 (497 mm SL), Ida, Numazu, Shizuoka, Japan, 250 m; HUMZ 185291, 185292 (2, 185– 258 mm SL), Hagi Bay, Yamaguchi, Japan, 10–20 m, mid-trawl; HUMZ 185353 (252 mm SL), Owase, Mie, Japan, 400 m; HUMZ 191786–191826 (41, 232– 820 mm SL), Kiinagashima, Mie, Japan, 150–300 m, bottom trawler; HUMZ 192387 (236 mm SL), Mito, Ibaraki, Japan, 310 m, bottom trawler Wakataka Maru; HUMZ 193074, 193075 (2, 395– 431 mm SL), fish market, Tashi, Taiwan.
Diagnosis. A species of Ateleopus with 8–10 dorsal-fin rays, 12–14 pectoral-fin rays, 96–131 anal-fin rays + caudal-fin rays, 0+8–11=8–11 gill rakers, two or three rows of small teeth on upper jaw, a large obtuse spine projecting from posterior margin of eye, tip of pelvic fin enlarged and round, and tail long (range 70–83% of SL, mean = 78%).
Description. Counts and measurements of Ateleopus japonicus , including the lectotype of A. japonicus designated in this study and the holotype and paratypes of A. tanabensis , are given in Table 1. Data for the lectotype (a stuffed specimen) and the other specimens are described separately.
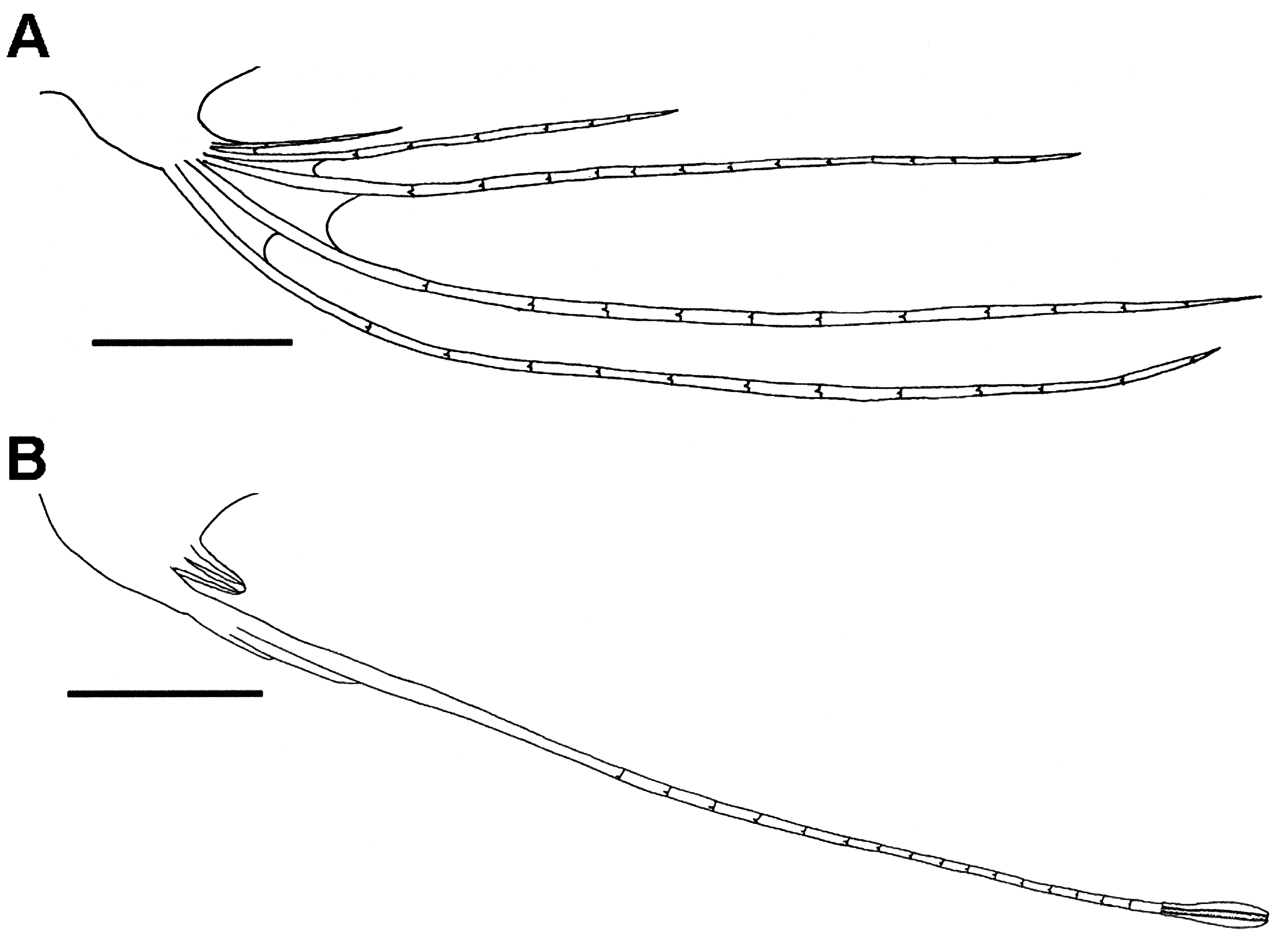
Description of lectotype. Dry mounted specimen. Body elongate, compressed; tail long, tapering to caudal fin; anus situated just in front of anal-fin origin; scales absent; trunk about 5.6 times in SL; body width at anal-fin origin about 1.6 times in head width. Head large, about 7.3 times in SL, slightly compressed, forming gelatinous snout, former gelatinous snout greatly reduced in size, its tip only reaching rostral most point of premaxilla. Nostrils open just in front of upper half of eye. Orbit small, left side with a glass eye, right side empty, four times in postorbital head length. Interorbital space wide with single shallow longitudinal depression. A large obtuse spine projecting laterally from posterior margin of eye; two small obtuse spines on each side between large obtuse spine and gill opening. Mouth inferior, protractile, rather small; upper jaw less than one-third of head length. Upper jaw with villiform tooth band; lower jaw with several villiform teeth in right jaw, arranged in one row and situated anteriorly; no teeth on left side. No teeth on palatine, vomer, or tongue. Symphysis of lower jaws convex in front. Gill opening wide; gill membranes separate, free from isthmus. Dorsal-fin origin slightly behind base of pectoral fin. Dorsal fin broken in half, the two parts barely connected. Dorsal-fin base short, less than snout length. Dorsal fin with 8 elongated rays, its height approximately equal to pectoral-fin length. Pectoral fin elongated, the tip (right side) extending to middle of trunk, its length longer than postorbital head length and shorter than head length without snout (anterior margin of orbit to posterior margin of operculum) tip of right side broken and barely connected; left pectoral-fin ray lacking posterior half. Precise count of pectoral-fin ray difficult because of damage and varnish covering. Pelvic fin jugular, appearing like one elongated ray, tip broken and barely connected, some anterior rudimentary rays attached to base of elongated ray, with a small cutaneous membrane on the medial side of the base; tip of longest ray slightly depressed and extending to anterior one-third of pectoral fin, right pelvic-fin ray lost from its base. Anal-fin base long, conspicuous over most of tail and continuous with caudal fin. Many tips of anal-fin rays damaged.
TABLE 1. Proportional measurements and counts for " Ateleopus japonicus ", " A. purpureus ", and " A. tanabensis " forms as described in text. Values in parentheses are means.
A. japonicus A. purpureus A. tanabensis
This study This study This study This study This study This study
Temminck lectotype of " A. japonicus" Tanaka " A. purpureus " holotype paratypes " A. tanabensis "
& Schlegel A. japonicus & (1915, 1916)
(1846) A. schlegelii
RMNH 1410 n=9 n=41 ZUMT 8320 n=3 n=19 Measurements
Total length (mm) 661 805+ 535–879 740 237–808 377 219–260 312–443 Standard length (mm) 800 500–820 715 227–764 345 212–243 281–405 % of SL
Pectoral-fin length 60.0 61.7–76.4 (70.1) 71.4–125.6 (91.8) 112.1 128.4–144.6 (139.1) 88.0–116.2 (101.3) Counts
Dorsal-fin rays 8 8 9– 10 9 9–10 10 9 9–10 Anal-fin rays 118 115+ 96–123 120 100–131 116 113–127 100–124 caudal-fin rays)
Pectoral-fin rays 12 11+ 12–14 13 12–14 13 12 12–14 rakers 0+8–11= 8–11 1 +8–9=9–10 0+8–11=8–11 0+10=10 0+9–10=9–10 0+9–11=9–11 Vertebrae 27–29+86–96=113–123 26–29+97–107=123–136 27+98=125 27+104=131 27–29+103–104=130–132
Color of mounted lectotype ( Fig. 2 View FIGURE 2 ). Left side and ventral region painted and varnished. Overall color on left side of head and dorsum medium blackish brown; ventral regions yellowish brown. Right side of head blackish brown dorsally, yellowish brown ventrally; body and tail brownish yellow. Dorsal and pectoral fins blackish brown; pelvic, anal and caudal fins yellowish brown.
Description of the other specimens. Body elongate, compressed; tail long, tapering to caudal fin, completely enveloped in soft gelatinous tissue, without scales except for thin scales embedded in lateral line. Head large, about five to ten times in SL, slightly compressed, covered with thick gelatinous tissue forming gelatinous snout in adults but not in juveniles (up to ca. 250 mm SL). Anterior nostril with a short tube; posterior nostril large, without a tube, situated just in front of upper half of eye. Eye small, its diameter slightly shorter than pectoral-fin base. Interorbital space wide, with a deep longitudinal groove. A large obtuse spine projecting from posterior margin of eye. Two small obtuse spines between large obtuse spine and gill opening. Mouth inferior, protractile, rather small. Posterior end of maxilla extending to, or a little beyond, center of eye. Both lips rough, densely covered with tiny tooth-like papillae. Upper jaw with one row of villiform teeth in specimens to ca. 320 mm SL, developing into a villiform tooth band in specimens larger than 320 mm SL; tooth band wider anteriorly; teeth in lower jaw small canines, very deciduous, arranged in one row and situated anteriorly; the lower jaw teeth sometimes all fallen out. Palatine, vomer, and tongue toothless. Symphysis of lower jaws convex in front. Preopercular margin not free; gill opening wide; gill membranes separate, free from isthmus. Upper margin of opercle usually acute, rarely rounded, protruding posteriorly. Uppermost gill raker of first gill arch, flattened, usually straddling the angle of arch (between epibranchial and ceratobranchial). Gill rakers on upper part of lower limb of first gill arch flattened, lower ones stumpy: rakers on other gill arches all stumpy. All tips of gill rakers bluntly pointed, the edge weakly spinulose. No pseudobranchiae. Branchiostegal rays usually seven. Dorsal-fin origin slightly behind base of pectoral fin. Dorsal-fin base short, less than snout length. Dorsal fin long, its length approximately equal to pectoral-fin length. Pectoral fin long, the tip not reaching origin of anal fin until a size of ca. 280 mm SL is attained; the tip reaching or extending beyond origin of anal fin in most of specimens from ca. 280 to ca. 385 mm SL, the tip not reaching origin of anal fin in the specimens larger than ca. 385 mm SL. Pelvic fin jugular, consisting of four autogenous rays in postlarva of 185 mm SL; four long rays and a minute ray in postlarva 258 mm SL ( Fig. 3 View FIGURE 3 A); in juvenile specimens ca. 227–270 mm SL anterior three long rays attached to each other, and cutaneous membrane enclosing posterior two rays, the third ray longest with swollen and slightly flattened tip ( Fig. 3 View FIGURE 3 B); in specimens larger than 270 mm SL anterior two rays short or rudimentary, attached to base of third ray, with vestigial rays in the cutaneous membrane. Precise count of pelvic-fin rays difficult in adults because of cutaneous membrane and rudimentary anterior two rays becoming very small. Proportional length of pelvic-fin ray variable, regardless of size, extending to a point anywhere between base of pectoral and proximity of anus. Anal-fin base long, continuous with caudal fin. Anal-fin pterygiophores visible through gelatinous tissue. Anus just in front of anal-fin origin.
Color when fresh. For postlarva: see Amaoka & Kobayashi (2003). Adult, as seen in Figure 4 View FIGURE 4 , blackish or dark purplish, translucent; all fins blackish except translucent first rays of dorsal and pectoral fins, and whitish tip of pelvic fin.
Color in preserved specimens. Postlarva uniformly milky-white on body. Adult brownish, blackish, grayish or translucent; all fins blackish except whitish first rays of dorsal and pectoral fins, and white tip of pelvic fin. Mitochondrial DNA analyses. An 810-bp region of the cytocrome b gene was successfully amplified by PCR and sequenced in all specimens selected. No ambiguous alignments were found, and no gaps had to be postulated.
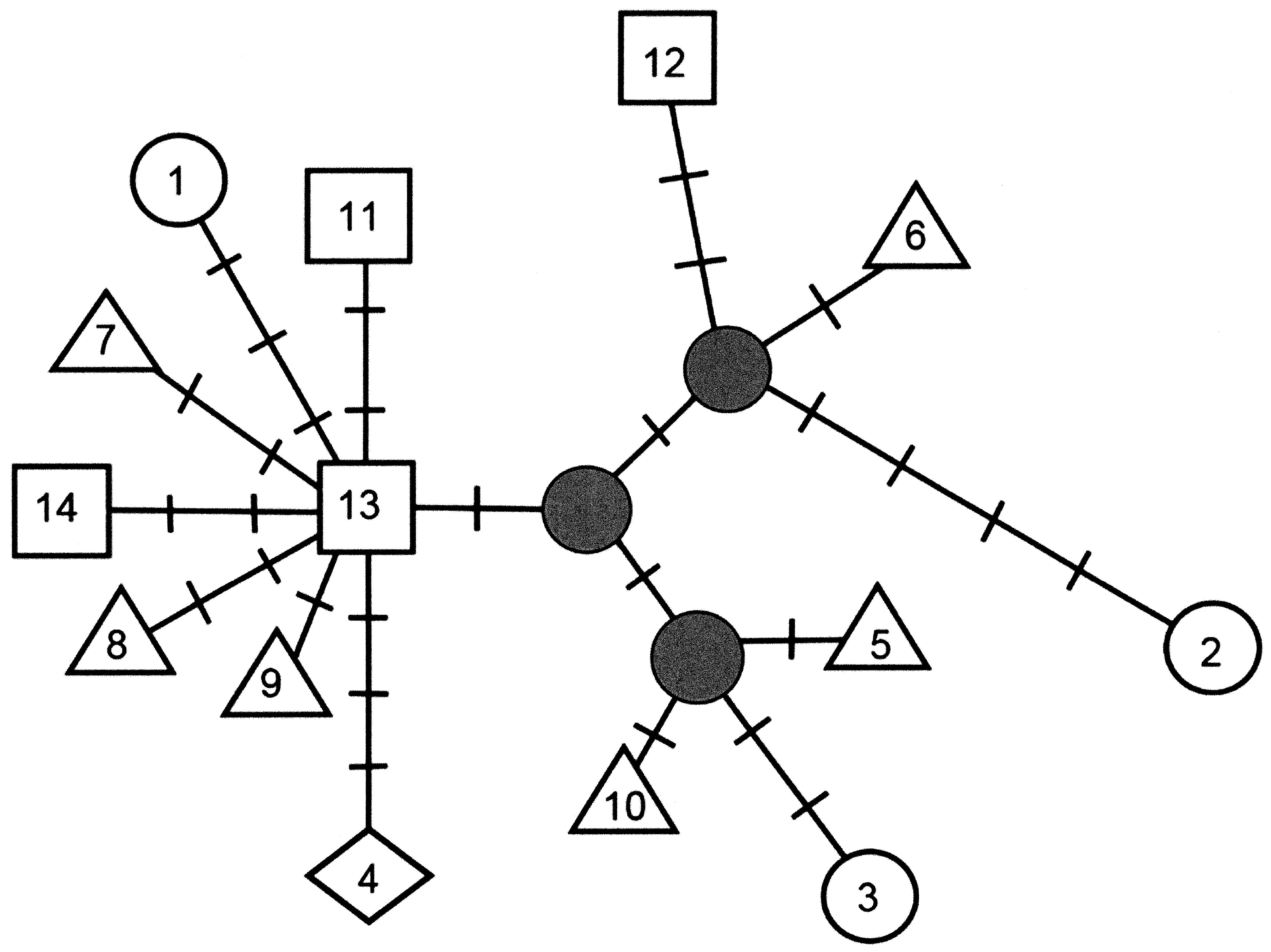
Twenty-four polymorphic sites were identified in the amplified region for the 17 specimens sequenced and downloaded, with 14 mtDNA haplotypes (1–14) identified (Table 2). “ Ateleopus japonicus ”, “ A. purpureus ”, and “ A. tanabensis ” were indicated as three morphotypes (short, medium, and long fin, respectively) as explained in discussion. The haplotypes 1–4, 6–12, and 14 are a single individual and the haplotypes 5 and 13 are suites of two and three individuals, respectively (Table 2). The voucher specimens of the sequences all were caught off Kiinagashima, Mie, Japan except for haplotype 4, 11–12 which are from Suruga Bay, Shizuoka, Japan. Haplotype 13 includes one specimen from Suruga Bay and two specimens from Kiinagashima. The voucher specimens for the sequences are represented among the specimens employed for morphological comparisons ( Fig. 9 View FIGURE 9 ), except for the haplotype 4 submitted by Miya et al. (2001). The voucher specimen of Ateleopus japonicus of Miya et al. (2001) is considered to be lost (M. Miya pers. comm.).
Pairwise sequence divergence among haplotypes was low-ranged from 0.124% (between Haplotype 9 and 13) to 1.122% (between Haplotype 1 and 2, and Haplotype 2 and 4) (Table 2).
A maximum parsimony network for the 14 haplotypes with 29 mutational steps and a consistency index of 0.90 was generated ( Fig. 5 View FIGURE 5 ).
TABLE 2. Pairwise nucleotide sequence divergence (%) among mtDNA cytochrome b haplotype in the three morphotypes (short, medium, and long fin) of the Ateleopus as described in text calculated by Kimura's 2 paramater method ( Kimura 1980)
Nucleotide sequence divergence (%)
Haplotype Specimens Morphotype 1 2 3 4 5 6 7 8 9 10 11 12 13 14 HUMZ 191790 short fin -
HUMZ 191796 short fin 1.122 -
HUMZ 191799 short fin 0.871 0.746 -
Miya et al. (2001) ( A. japonicus ) 0.621 1.122 0.871 -
HUMZ 191788, 191794 medium fin 0.745 0.872 0.372 0.745 -
HUMZ 191795 medium fin 0.496 0.621 0.621 0.745 0.496 -
HUMZ 191791 medium fin 0.620 0.998 0.746 0.620 0.621 0.621 -
HUMZ 191797 medium fin 0.620 0.996 0.745 0.620 0.620 0.620 0.496 - HUMZ 191798 medium fin 0.496 0.872 0.621 0.496 0.496 0.796 0.372 0.371 - HUMZ 191800 medium fin 0.745 0.872 0.372 0.745 0.248 0.496 0.621 0.620 0.496 - FKUN 33359 long fin 0.620 0.998 0.746 0.620 0.621 0.621 0.496 0.496 0.372 0.621 - FKUN 33361 long fin 0.870 0.745 0.745 0.870 0.620 0.371 0.745 0.745 0.620 0.620 0.745 - FKUN 33360, HUMZ 191789, 0.371 0.746 0.496 0.371 0.372 0.372 0.248 0.247 0.124 0.372 0.248 0.496 - 191793 long fin
HUMZ 191792 long fin 0.620 0.745 0.496 0.620 0.620 0.620 0.496 0.496 0.247 0.620 0.496 0.745 0.247 - Distribution. Known from along coasts of Japan northward to southern Tohoku district, Okinawa Trough; East and South China Seas; north-western Western Australia and eastern Australia; Chesterfield Island in Coral Sea. Depth range from 100 to 600 m, on sandy-muddy bottoms.







No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Ateleopus japonicus Bleeker 1853
| Kaga, Tatsuya, Van Oijen, Martien J. P., Kubo, Yoshikazu & Kitagawa, Emi 2015 |
Ateleopus tanabensis
| Aizawa 2013: 410 |
| Shinohara 2009: 701 |
| Kitagawa 2008: 28 |
| Aizawa 2002: 347 |
| Mochizuki 2001: 109 |
| Shinohara 2001: 299 |
| Randall 2000: 591 |
| Matsubara 1955: 273 |
| Okada 1938: 62 |
| Howell 1935: 96 |
| Tanaka 1918: 223 |
Ateleopus purpureus
| Aizawa 2013: 410 |
| Shinohara 2009: 701 |
| Prokofiev 2006: 342 |
| Shinohara 2005: 407 |
| Aizawa 2002: 347 |
| Shinohara 2001: 299 |
| Xu 1988: 145 |
| Okamura 1985: 439 |
| Matsubara 1955: 273 |
| Okada 1938: 62 |
| Howell 1935: 96 |
| Tanaka 1916: 396 |
| Tanaka 1915: 565 |
Ateleopus schlegelii
| Hoeven 1855: 326 |
Ateleopus japonicus
| Okiyama 2014: 269 |
| Aizawa 2013: 410 |
| Fricke 2011: 358 |
| Shinohara 2009: 700 |
| Bray 2006: 457 |
| Shinohara 2005: 407 |
| Amaoka 2003: 109 |
| Aizawa 2002: 347 |
| Mochizuki 2001: 109 |
| Shinohara 2001: 298 |
| Randall 2000: 591 |
| Kulbicki 1994: 14 |
| Mochizuki 1984: 115 |
| Shen 1972: 24 |
| Matsubara 1955: 272 |
| Okada 1938: 62 |
| Howell 1935: 93 |
| Sauter 1905: 234 |
| Bleeker 1853: 19 |
| Temminck 1846: 255 |