
Neoromicia isabella, Decher & Hoffmann & Schaer & N Orris & Kadjo & Astrin & Monadjem & Hutterer, 2015
|
publication ID |
https://doi.org/10.3161/15081109ACC2015.17.2.003 |
|
DOI |
https://doi.org/10.5281/zenodo.4335962 |
|
persistent identifier |
https://treatment.plazi.org/id/039C0121-FFF7-FFCE-761B-FBD71C18554D |
|
treatment provided by |
Felipe |
|
scientific name |
Neoromicia isabella |
| status |
sp. nov. |
Neoromicia isabella View in CoL View at ENA sp. nov. Decher, Hutterer and Monadjem
Isabelline White-winged Serotine
Neoromicia cf. rendalli View in CoL ; Monadjem et al., 2013 b
Holotype
ZFMK 2008.0292 About ZFMK , field number JD 650, collected by Jan Decher on 7 March 2008. Adult male in reproductive condition (testes 5 × 4 mm), preserved in 70% ethanol, skull extracted. Tissue preserved (COI sequence GenBank no. KT598187 View Materials ). View Materials
Type locality
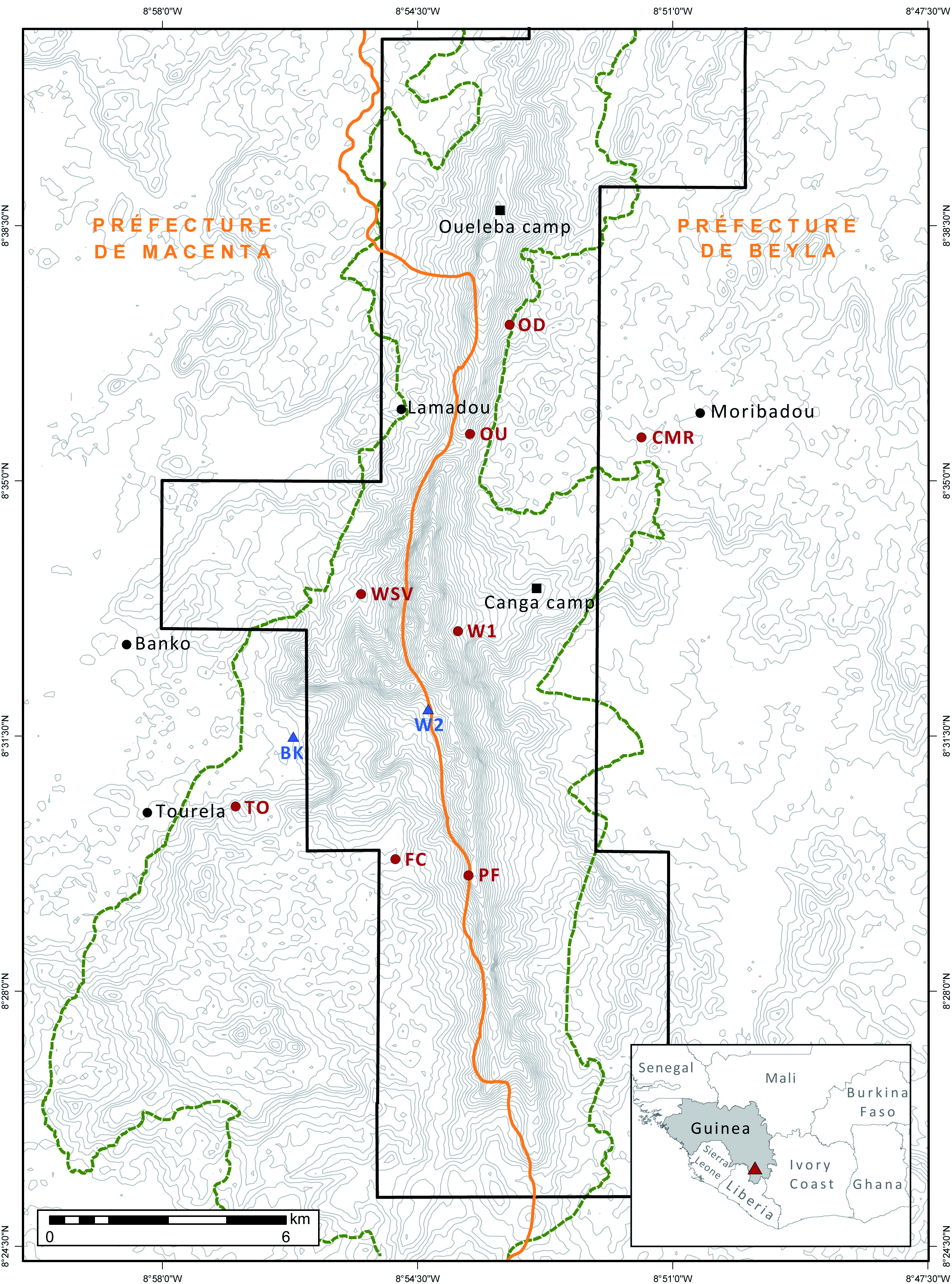
Guinea, Province Macenta, Simandou Range, Foko Confluence, 8°29’48.62’’N, 8°54’48.22’’W, ca. 765 m a.s.l., captured in net set across creek in rainforest.
Paratype
ZFMK 2008.0291 About ZFMK , field number JD 647 ; adult female (pregnant with 1 embryo), preserved in ethanol. Netted same day and place as holotype.
Referred specimen
DM 12619 , Mount Nimba, Liberia ; reported as Neoromicia cf. rendalli by Monadjem et al. (2013 b); COI sequence deposited under GenBank no. JX508832 View Materials .
Diagnosis
A medium-sized pipistrelloid bat of the genus Neoromicia , based on the presence of a single upper premolar (Hill and Harrison, 1987) with white wings; only likely to be confused with N. tenuipinnis (slightly smaller and different pelage colour, see below) and N. rendalli (larger). Dorsal and ventral surface of wings pure white (as in N. tenuipinnis ), tips of dorsal pelage light rusty orange-brown (‘isabelline’), not dark brown (as in N. tenuipinnis ). Baculum similar in shape to N. rendalli but smaller. COI sequence different from that of both N. tenuipinnis and N. rendalli ( Fig. 3 View FIG ).
Etymology
The species epithet refers (as a noun in apposition) to the ‘Isabella’ colouration (sensu Ridgeway, 1912) of the dorsal fur of this bat, and also recalls the name of Jan Decher’s daughter Isabelle.
Description
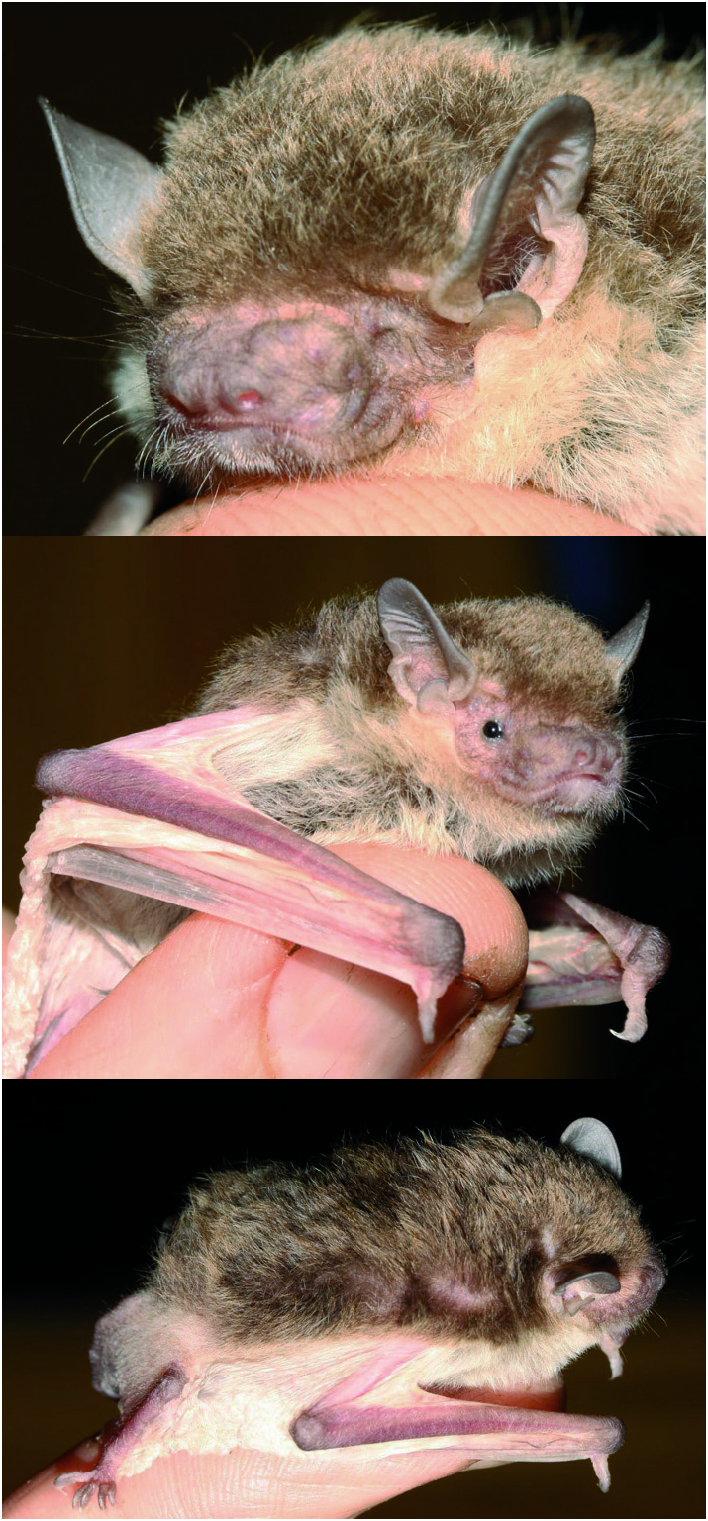
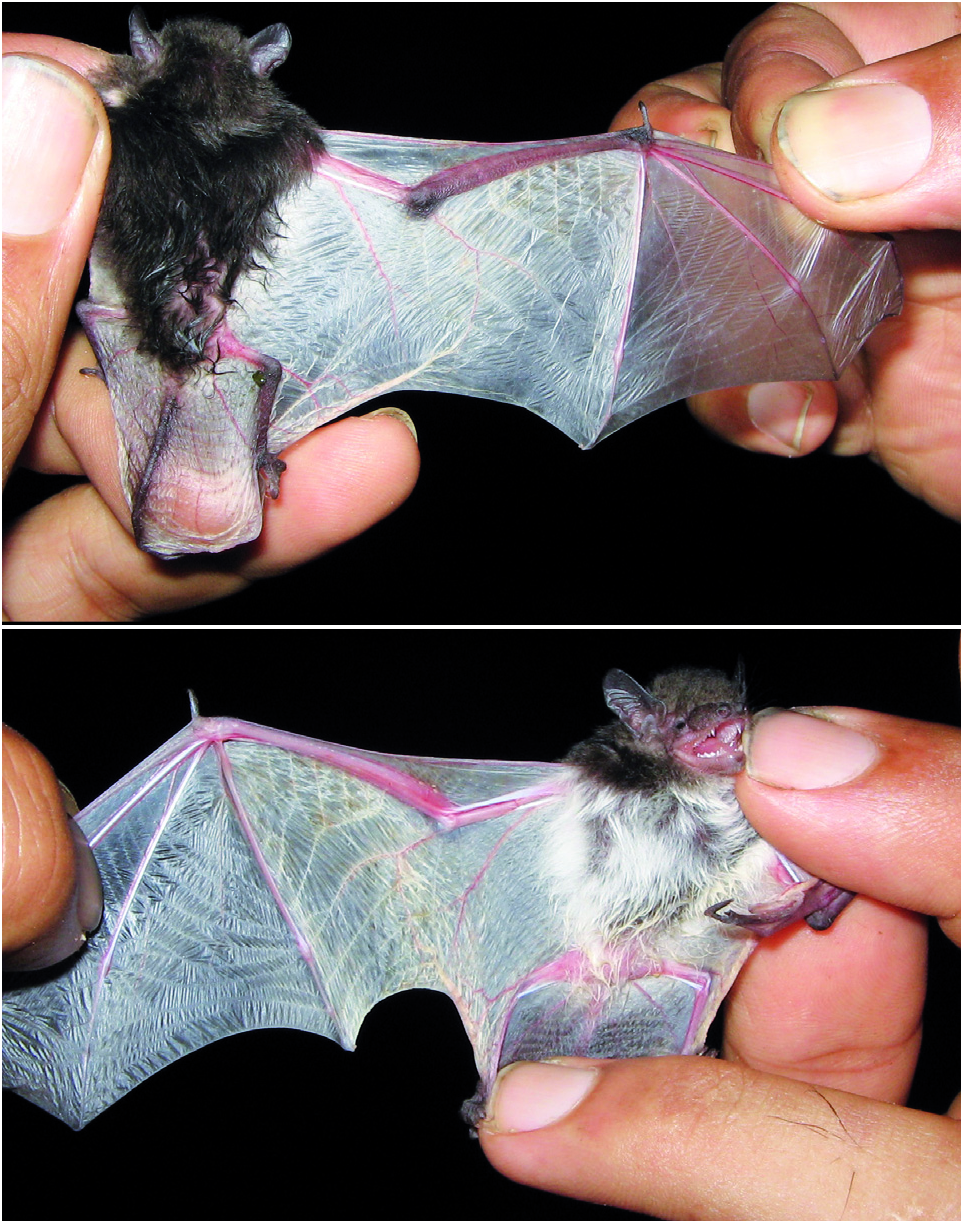
Neoromicia isabella sp. nov. is similar in size and general colour pattern to N. tenuipinnis ( Table 3 View TABLE , Figs. 4 View FIG and 5 View FIG ) but slightly larger in total length, forearm length, and body mass ( Table 3 View TABLE ). Tail length is 34% of the total length in the holotype, and 37% on average. This value in the holotype of N. rendalli ( Thomas, 1889) is 49%, and in the holotype of N. tenuipinnis 39% ( Peters, 1872). The tips of the dorsal hairs are of a light isabelline colour ( Ridgeway, 1912), much lighter than in N. tenuipinnis (compare Figs. 4 View FIG and 5 View FIG ). The dorsal hairs are 4.5 mm long. Ears, lips, wings, uropatagium, and hindfeet are almost white and covered by very short hairs; only the ear tips and dorsal surfaces of forearm, hindfeet and tail have a pale brownish hue.
Ear length is 10 mm in the holotype of N. isabella ( 10–13 mm, n = 3), and its tip is rounded ( Fig. 4 View FIG ). The tragus of the holotype ( Fig. 6 View FIG ) is 4.2 mm long, and represents 42% of the ear length. A small notch is present at the basis of the anterior surface. Compared to illustrations of N. tenuipinnis and N. rendalli in Rosevear (1965) and specimens in ZFMK and DM, the tragus of N. isabella is similar to that of N. tenuipinnis (tragus length estimated from figure: 3.9 mm) but the shape of the notch is different ( Fig. 6 View FIG ).
The penis of the holotype specimen is 6.4 mm long and covered by short whitish hairs. A digital x -ray of its baculum is shown in Fig. 7A View FIG in a semilateral view. It is slightly longer ( 3.5 mm) than in the N. isabella specimen DM12619 ( 2.8 mm; Fig. 7B View FIG ) reported by Monadjem et al. (2013 b: Fig. 4C View FIG ), but very similar in shape, and both are still longer than that of N. tenuipinnis (in Monadjem et al., 2013 b: Fig. 4D View FIG ), which is 1.8 mm long and strongly bent ( Fig. 7C View FIG ). A baculum figured as N. rendalli by Kearney et al. (2002: Fig. 1F View FIG ) is about 4 mm long.
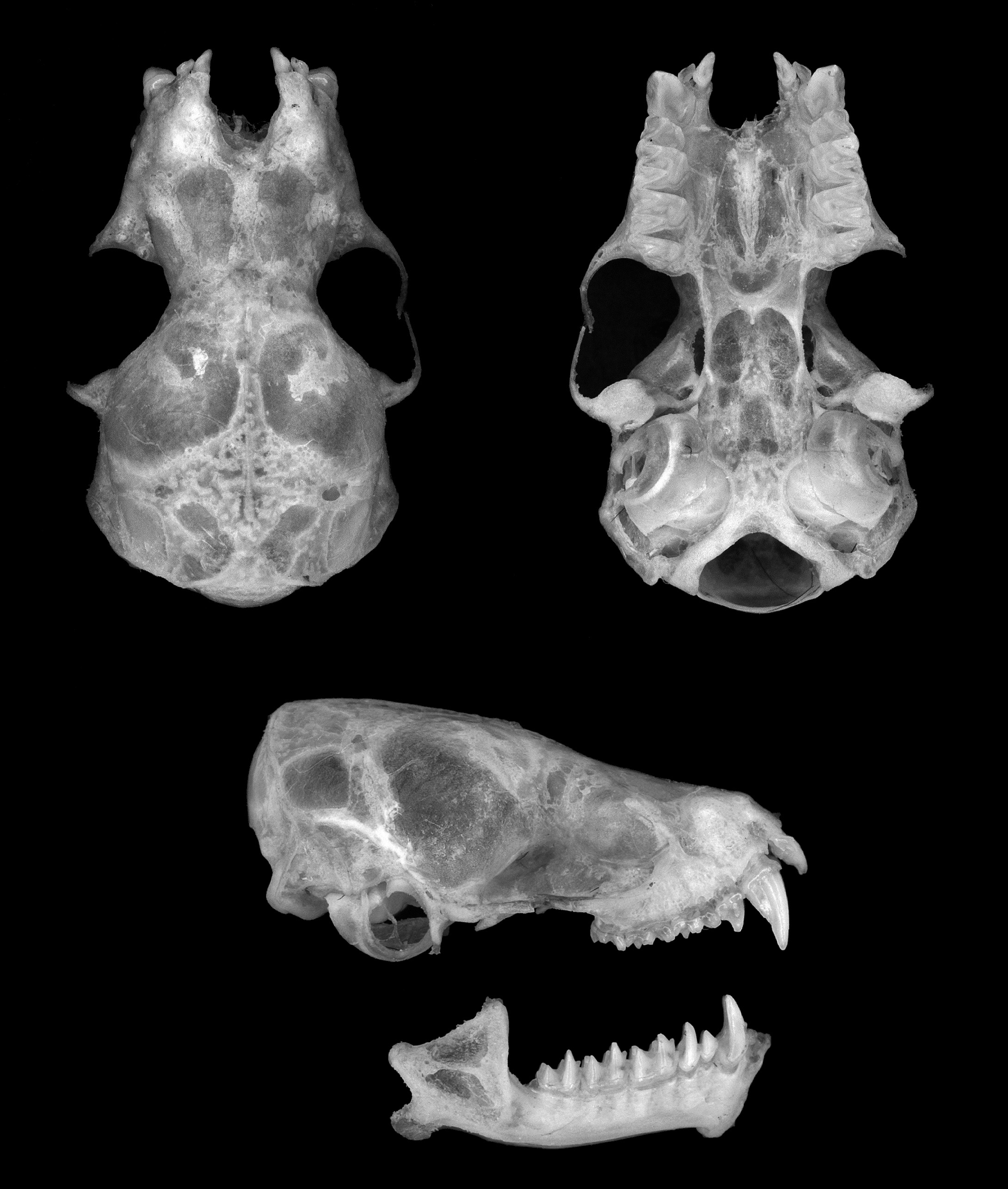
The cranium of N. isabella is illustrated in Fig 8 View FIG . Neoromicia isabella is distinguished from N. tenuipinnis (skull figured in Monadjem et al., 2010: 479) by slightly larger cranial and dental measurements ( Table 4 View TABLE ), particularly in the greatest skull length and the upper and lower molar rows. Greatest skull length in N. rendalli (skull figured in Monadjem et al., 2010: 475) is similar to that of N. isabella , but its cranium is wider, as expressed by zygomatic width, greatest width of braincase, and mastoid width ( Table 4 View TABLE ).
Within the genus Neoromicia the greatest skull length of N. isabella (GSKL 12.5–13.0 mm) is of medium size and less robust than in N. rendalli , with a narrow maxillary and braincase and a fragile zygomatic arch. Skulls of 15 other species assignable to the genus Neoromicia (see accounts in Monadjem et al., 2010, 2013 b; Goodman et al., 2012; Happold and Happold, 2013) are either larger or smaller in length. The dorsal profile of the skull is slightly concave at the rostrum and slopes gradually upwards to the occiput. There is no sagittal crest, but a faint lambdoid crest. The dental formula is I2/3, C1/1, P1/2, M3/3 = 32, as in the other Neoromicia . Several observers have reported the presence of a tiny anterior upper premolar present on either or both sides of the upper jaw in N. tenuipinnis ( Hayman, 1954; Schlitter et al., 1982; Thorn and Kerbis Peterhans, 2009). The occasional presence of this tiny anterior upper premolar is not unusual in Neoromicia species (e.g. see Rosevear, 1965; Happold and Happold, 2013); however, not one of the specimens that we examined displayed such a tooth. The anterior upper incisor is more than twice as long as the posterior (about twice in N. rendalli and N. tenuipinnis — Monadjem et al., 2010) and unicuspid. The upper canine is long; 1.6-times as long as the anterior upper incisor, and 2.5-times as long as the subsequent premolar. The upper molar rows (C–M 3) are almost parallel; the molars are shorter than wide, and all teeth from the canine to the third molar are in contact. The mandible length is intermediate between that of N. tenuipinnis and N. rendalli ( Table 4 View TABLE ). The lower incisors are all trifid.
Comparisons and Designation of a Neotype for Neoromicia tenuipinnis
Our comparisons are hampered because the holotype of N. tenuipinnis is lost (Turni and Kock, 2008), and the original description of Peters (1872) does not include skull measurements. Likewise, the description of N. rendalli by Thomas (1889) does not contain skull measurements; however, the holotype skull (BMNH 1889.3.2.3) is figured in Monadjem et al. (2010) and in the ACR (2015: Appendix), and was examined by AM. External measurements of the types of both taxa, however, are congruent with the current treatment of the species ( Table 3 View TABLE ).
Externally the new species is quite distinct. Our images ( Fig. 4 View FIG ) may be compared with those of N. tenuipinnis from Liberia ( Fig. 5 View FIG ) and another one from Comoe National Park, Côte d’Ivoire (http:// www.inaturalist.org/photos/189398). The latter species has much darker body coloration. Neoromica rendalli is also darker, and the dark-brown coloration extends to the ventral surface of the body (specimen ZFMK 1977.1015 from South Sudan). Neoromicia isabella is immediately distinguishable from its sister species N. roseveari , itself only recently described, by its white wings and the rich colour of its fur. In N. roseveari , the wings are dark and the fur is dark brown ( Monadjem et al., 2013 b). Furthermore, N. roseveari is far larger (for the holotype, FA is 37.1 mm and GSKL is 14.36 mm).
The whole group, however, may include further unrecognized taxa if one looks at the genetic and morphological variation throughout Africa, which is beyond the scope of our paper. Specimens from Nigeria to Central Africa mentioned by Bergmans (1977), Viellard (1974), Thorn and Kerbis Peterhans (2009), Bates et al. (2013), and other authors should be re-examined in the context of a revision of the genus. It should be also noted that the most current range map for N. tenuipinnis sensu lato ( Fahr, 2013 e) may contain records of both N. tenuipinnis sensu stricto (see below) and the newly described N. isabella .
Under these circumstances, however, we consider it essential to fix the morphological and genetic characters of N. tenuipinnis and to designate a neotype for Vesperus tenuipinnis Peters, 1872 (now Neoromicia tenuipinnis ), because the former holotype, an adult female from ‘Guinea’ ( Peters, 1872: 264), in the Berlin Museum (ZMB) is lost (Turni and Kock, 2008). Monadjem et al. (2013 b) listed a specimen in the Natural History Museum, London (BMNH 1889.5.1.3) as a ‘syntype’ of N. tenuipinnis , which however is incorrect, as Peters (1872: 264) only mentioned a single specimen in the Berlin Museum at hand; the one, which is now lost.
Here we designate specimen DM 13235 in the Durban Natural Science Museum as the neotype. It consists of a male specimen collected by A. Monadjem at Bonlah village , near Yekepa at the base of Mount Nimba, Liberia on 7 January 2012; external and cranial measurements are included in Tables 3 View TABLE and 4 View TABLE . The body is preserved in formalin with the skull extracted. The tragus and the baculum (also Fig. 7C View FIG ) are documented in Monadjem et al. (2013 b: Figs. 4D View FIG and 6D View FIG ). A COI sequence is deposited in GenBank ( JX508831 View Materials ). The external measurements of the neotype are very similar to those of Peters’ (1872) lost holotype ( Table 3 View TABLE ).
In light of our new genetic analysis ( Fig. 3 View FIG ), the genus Neoromicia , at least in West Africa, no longer appears to be paraphyletic (as suggested by Monadjem et al., 2013 b) but consists of a monophyletic clade. The type species of the genus, N. zuluensis (Roberts, 1924) , was studied genetically by Goodman et al. (2012) showing that it is sister to N. somalica . We are therefore confident that our new species has been assigned to the correct genus. In our COI tree N. isabella is sister to N. roseveari , and both taxa sister to N. brunnea , while N. tenuipinnis and N. rendalli group more at the root of this clade ( Fig. 3 View FIG ). Hence, whereas morphological characters suggest a closer relationship between N. tenuipinnis and N. isabella , this is not supported by the genetic analysis.
Remarks
This is the first record of a white-winged bat from the Simandou Range, initially identified as N. tenuipinnis . Eleven individuals were captured, all between 7 and 9 March 2008 in the canopy net over the creek at FC, two of which were kept as vouchers. They group with specimen DM12619 from Mount Nimba, Liberia, that was labelled Neoromicia cf. rendalli by Monadjem et al. (2013 b). Neoromica isabella was not recorded during the 2002 RAP, nor the 2003 RAP. The individuals captured in the context of this study over the forest creek at FC suggests that N. isabella uses drainages lines as travel routes probably emerging from hollow trees or rock crevices upstream. The specimen from the Liberian portion of Mount Nimba was captured emerging from the roof of a hut in a small village close to the larger settlement of Yekepa ( Monadjem et al., 2013 b).
Records from adjacent regions ( Aellen, 1963; De Vree, 1971; Bützler, 1994; Fahr et al., 2006; Monadjem and Fahr, 2007) may refer either to N. tenuipinnis or to the new species. Monadjem et al. (2013 b) collected both species on Mount Nimba. A specimen mentioned by Roche (1971) as N. tenuipinnis deviates by very small external and cranial measurements, and until a re-examination of the voucher specimen we regard this record with caution.
Conservation status
The conservation status of this species has not yet been evaluated ( IUCN, 2015).



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Neoromicia isabella
| Decher, Jan, Hoffmann, Anke, Schaer, Juliane, N Orris, Ryan W., Kadjo, Blaise, Astrin, Jonas, Monadjem, Ara & Hutterer, Rainer 2015 |
Neoromicia cf. rendalli
| Monadjem et al. , 2013 b |