
Poecilimon pseudornatus, Ingrisch, Sigfrid & Pavićević, Dragan, 2010
|
publication ID |
https://doi.org/10.5281/zenodo.197230 |
|
DOI |
https://doi.org/10.5281/zenodo.6204482 |
|
persistent identifier |
https://treatment.plazi.org/id/039D4825-F652-FFF0-FF12-FC7AFD42FA46 |
|
treatment provided by |
Plazi |
|
scientific name |
Poecilimon pseudornatus |
| status |
sp. nov. |
Poecilimon pseudornatus sp. n.
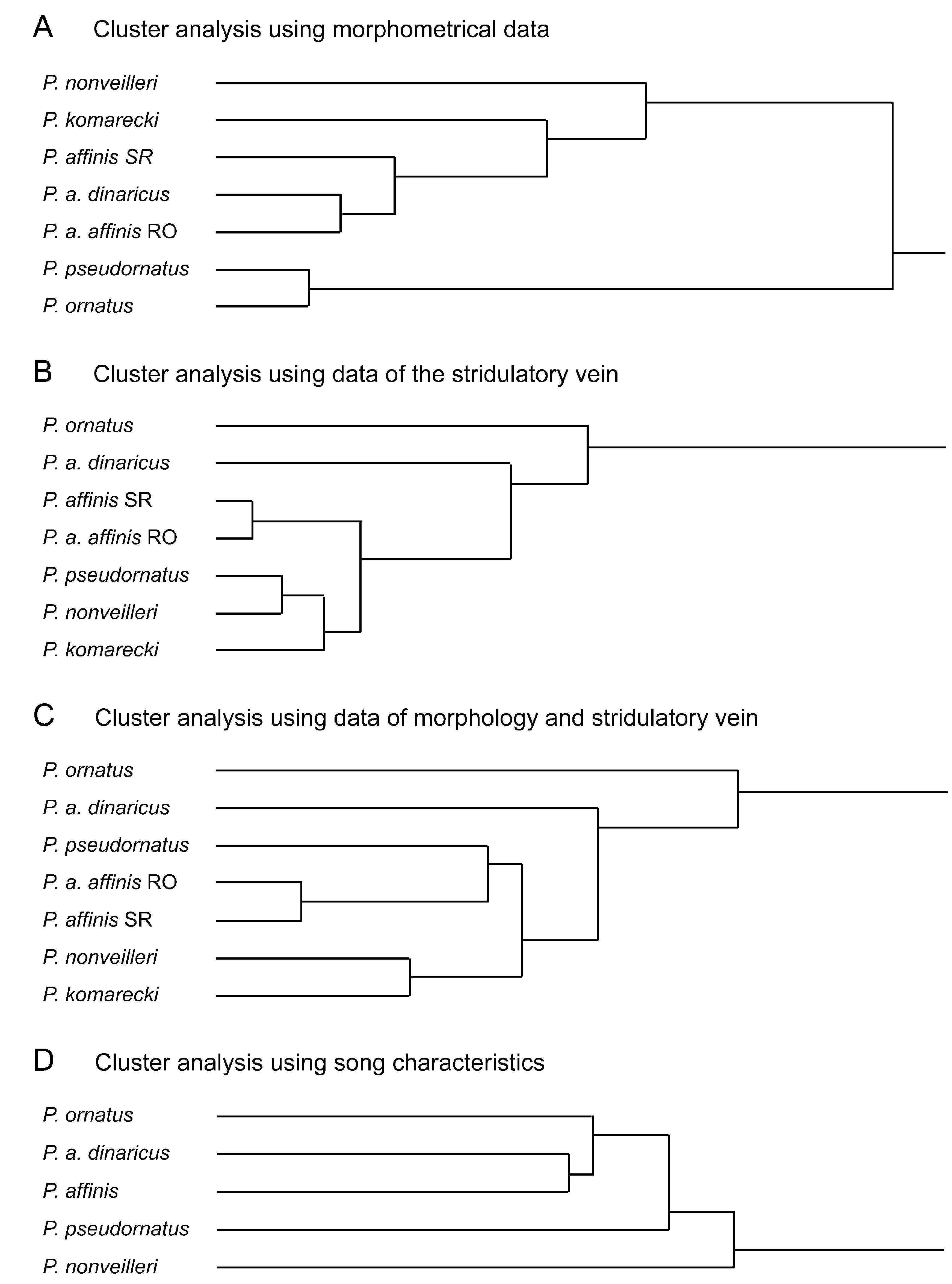
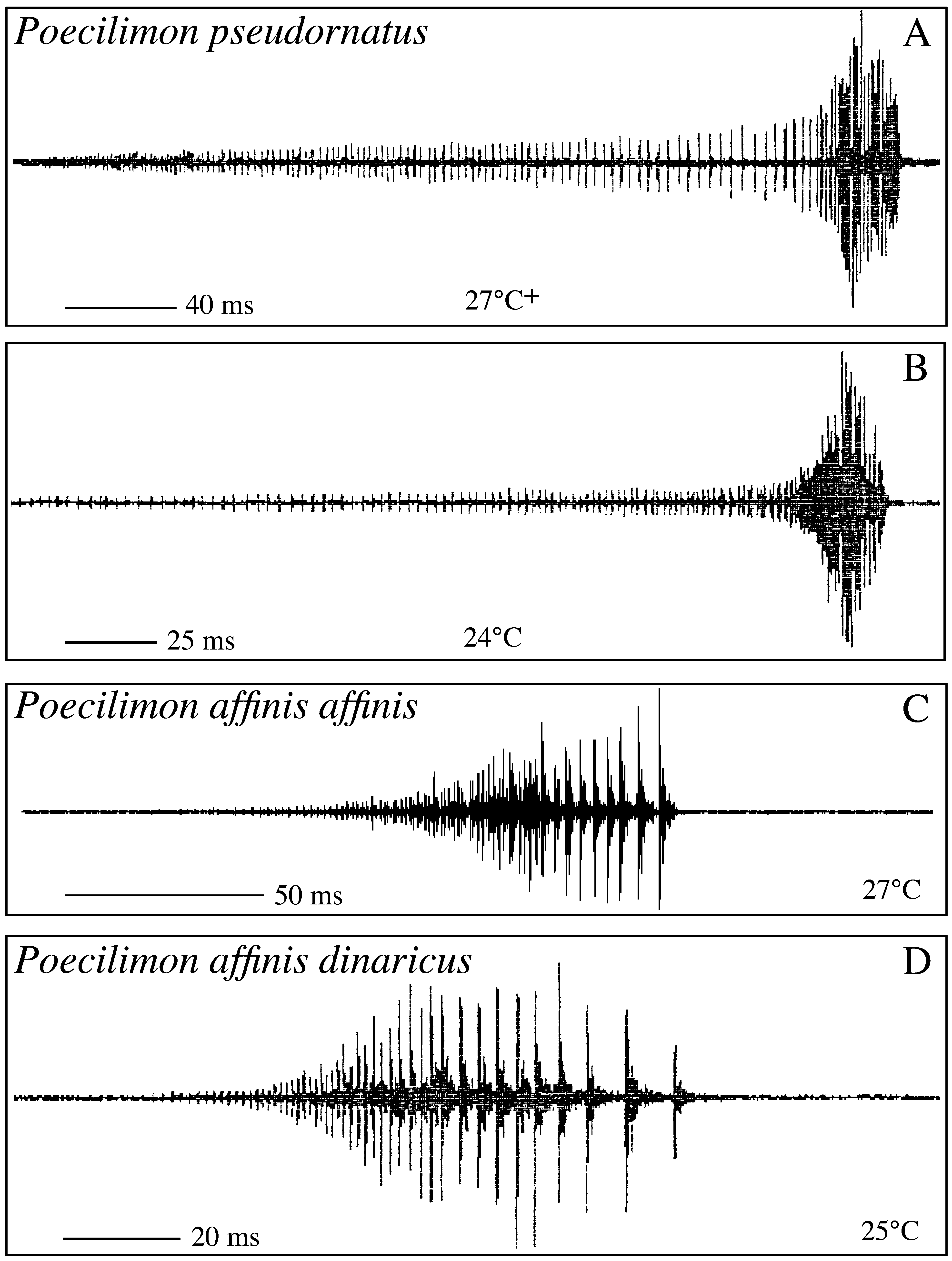
Figs. 6 View FIGURE 6 , 7A–H View FIGURE 7. A – H , 9 View FIGURE 9 A, 11A–B; Tabs. 5–6 View TABLE 5 View TABLE 6
Holotype (male): Crna Gora [ Montenegro], Durmitor, Donji Unač, 1200–1300m, 29.VII.1988, in Coll. Muséum d'Histoire naturelle de Genève ( MHNG).
Paratypes: Montenegro, Durmitor: 17 3, 6 Ƥ [including allotype], Donji Unač, 1200–1300m, 29.VII.1988 + 5.VIII.1988, leg. Ingrisch & Pavićević (allotype in MHNG, remainder in CI + CDPV); 13 3, 6 Ƥ, Crno jezero, Čeline, 1500m, 6.VIII.–10.VIII.1985, leg. Ingrisch & Pavićević (CI + CDPV); 9 3, 5 Ƥ, do., 20.VII.1987; 1 3, 1 Ƥ, do., 27.VII.1988; 3 3, 2 Ƥ, do., 14.VIII.1989; 1 Ƥ, same locality, 22.VII.1991, leg. D. Pavićević ( CDPV); 3 3, 7 Ƥ, do., 10.VIII.1992; 1 3, do., 4.VIII.1993; 1 3, 1 Ƥ, Tepca, 1000m, 21.VII.1985, leg. D. Pavićević ( CDPV); 4 3, 1 Ƥ, do., 27.VI.1988; 2 3, same locality, 1.VIII.1988, leg. S. Ingrisch (CI); 1 3, Petrovački Potok, 1250–1400m, 21.VII.1987, leg. S. Ingrisch (CI); 1 3, Tepca, 500–1100m, 1.VIII.1988, leg. S. Ingrisch (CI); 1 3, Trsa, 1400m, 5.VII.1988, leg. D. Pavićević ( CDPV); 10 3, 3 Ƥ, Trsa, 1200–1400m, 4. VIII–6.VIII.1989, leg. Ingrisch & Pavićević (CI + CDPV); 3 3, 3 Ƥ, Dubrovsko-Bezuje, 1300–1400m, 10.VIII.1989, leg. D. Pavićević ( CDPV); 1 3, Pošćenski kraj, Virak, 1500m, 5.VIII.1989, leg. D. Pavićević ( CDPV); 1 3, Jarčište, 1500m, 7.VIII.1990, leg. S. Ingrisch (CI); 1 3, Podmilogora-Ninkovići, 1400–1500m, leg. D. Pavićević ( CDPV);1 3, 2 Ƥ, Pirlitor, Pleća, 1000m, 2.VII.1990, leg. D. Pavićević ( CDPV); 5 3, 1 Ƥ, Pirlitor, Seline, 1000–1100m, 5.VIII.1990, leg. Ingrisch & Pavićević (CI + CDPV); 8 3, 3 Ƥ, same locality ex ovo, leg. S. Ingrisch (CI); 1 3, same locality, 2.VIII.1991, leg. D. Pavićević ( CDPV); 1 3, do., 7.VIII.1992; 1 3, Pirlitor, 1400m, 2.VIII.1991, leg. D. Pavićević ( CDPV); 1 3, Seljkovac, 1600m, 7.VIII.1990, leg. D. Pavićević ( CDPV); 3 3, 2 Ƥ, Bjelasica Mt., Biogradska gora, 900m, 4.VII.1990, leg. D. Pavićević ( CDPV); 1 3, do., 7.VII.1991.
Other material studied: Serbia, 4 3, 1 Ƥ, Tara Mt., Mitrovac, 1080m, 20.VIII.1982, leg. I. Toševski ( CDPV); 24 3, 9 Ƥ, Tara Mt., Zaovine, 1000–1200m, 4.VIII.1989, leg. Ingrisch & Pavićević (CI + CDPV); 2 3, Tara Mt., Perućac, 12.VII.1990, leg. N. Ilić ( CDPV); 1 3, 1 Ƥ, Tara Mt., Kaluđerske Bare, 1000m, 12.VII.1990, leg. N. Ilić ( CDPV); 1 3, Povljen Mt., Debelo Brdo, 1000m, 3.VII.1990, leg. S. Ognjenović ( CDPV); 1 Ƥ, Murtenica Mt., 1200m, 10.VIII.1991, leg. Z. Bosiljčić ( CDPV); 5 3, Zlatibor Mt., Tekma, 1000m, 15.VIII.1995, leg. D. Pavićević ( CDPV); 3 3, 1 Ƥ, Pešter Plateau, Krstača, 1500m, 29.VII.2008, leg. P. L az a re vi ć ( CDPV); 2 3, Boljare, 1200–1300m, 30.VII.2008, leg. D. Pavićević ( CDPV); 1 3, 1 Ƥ, Kamena Gora (between Prijepolje and Brodarevo), 1100m, 16.VII.2008, leg. P. Lazarević ( CDPV); 1 3, Kamena Gora, 1100m, Ulica, 27.VIII.2008, leg. D. Pavićević ( CDPV).
Type locality. Montenegro, Durmitor, forest meadow within beech forest at the road Trsa–Piva below Donji Unač.
Measurements. see Tabs. 5–6 View TABLE 5 View TABLE 6 .
Diagnosis. The new species is similar to P. o r n a t u s. It mainly differs by the male stridulatory file and stridulation and by the longer ovipositor as outlined in the key and discussed above.
Description. A rather large, moderately stout species. Fastigium verticis ( Fig. 7E View FIGURE 7. A – H ) broader than scapus, or exceptionally only as broad as scapus, shallowly furrowed above. Postfemora with 4–13 spinules on interior and exterior ventral margins.
Male. Pronotum ( Fig. 7A–D View FIGURE 7. A – H ) moderately to distinctly raised in metazona (in a few males bred in laboratory ex ovo strongly raised), lateral margins slightly diverging in metazona; apex faintly concave to almost truncate. Tegmina usually covered by pronotum for circa half of their length. Stridulatory file ( Fig. 9 View FIGURE 9 A) with 187–241 stridulatory teeth ( Tab. 6 View TABLE 6 ), of which 110–160 are narrow. Cerci ( Fig. 7F–G View FIGURE 7. A – H ) curved in apical quarter but less strongly than in the following species, gradually narrowing into apical tooth. Subgenital plate with lateral margins converging in apical half, with a median keel in apical half distinct or only indicated; apex truncate to faintly excised.
Female. Pronotum faintly raised in metazona; apex truncate. Tegmina overlapping, largely hidden under pronotum; protruding part rounded. Cerci slightly curved, apex subacute. Subgenital plate small with a weak median keel; apex rounded to almost triangular, with or without a small median emargination. Ovipositor ( Fig. 7H View FIGURE 7. A – H ) long, apex dentate.
Coloration. Antennae annulated in all colour variants and in both sexes.
Male. Light form: Green with small black dots. Discus of pronotum partly yellow, especially in apical half; metazona with two red lateral cuneate spots, sometimes fused along apical margin. Tegmina yellow, discus infumate with black or brown. Abdominal tergites green with two white dorso-lateral bands.
Medium form: Yellow and red coloration on pronotum more pronounced. Abdominal tergites with four (or only two) black cuneate spots at the base of some or all tergites on both sides of the white bands. The black spots can be fused to one median and two lateral, ± continuous black bands.
Dark form: Yellowish brown to olivaceous green, dotted with rather large black spots. Vertex and dorsal area of femora and tibiae black. Pronotum with a median and two lateral black bands, also black along sulcus and apical margin. Abdominal tergites with a median and two lateral black bands, last tergite almost completely black. Epiproct, cerci, and subgenital plate yellow.
Female. Light form: Green with indication of two red cuneate spots in metazona of pronotum. Tegmina yellow.
Medium form: The red cuneate spots more distinct. Transverse black spots at the base of some abdominal tergites.
Dark form: As in male.
Etymology. Named for the similarity with P. ornatus .
Distribution. With certainty only known from the mountains of Montenegro (Durmitor) and southwest Serbia (Tara Mt., Zlatibor Mt., Kamena Gora between Prijepolje and Brodarevo, Pešter Plateau), where it lives at elevations of about NN+ 1000–1500m. We also found similar forms in Macedonia and south-eastern Serbia. The taxonomic status of those populations can however only be settled after extensive studies of stridulation.
Stridulation ( Fig. 11 View FIGURE 11 A–B): Males start singing in the morning, in laboratory they were also singing during darkness in the evening. Stridulation is monosyllabic. A prolonged, faint "sssss" is terminating in a sudden, short, loud "zip". The main frequency is between 14–16 kHz. Often a variable number of syllables are repeated in short intervals to form loose sequences.


| MHNG |
Museum d'Histoire Naturelle |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Phaneropterinae |
|
Genus |