
Chorizopora rosaria, Souto, Javier, Reverter-Gil, Oscar & Ostrovsky, Andrew N., 2014
|
publication ID |
https://doi.org/10.11646/zootaxa.3795.2.3 |
|
publication LSID |
lsid:zoobank.org:pub:6DE4B4A5-9369-4F9A-9F1A-C61EC4528F3A |
|
DOI |
https://doi.org/10.5281/zenodo.6144308 |
|
persistent identifier |
https://treatment.plazi.org/id/039D879A-FFA7-4C6C-FF16-FBE0FC7EFDE9 |
|
treatment provided by |
Plazi |
|
scientific name |
Chorizopora rosaria |
| status |
sp. nov. |
Chorizopora rosaria n. sp.
( Figs 15–20 View FIGURES 15 – 21. 15 ; Table 3 View TABLE 3 )
Chorizopora annulata (Lamouroux) : Norman, 1909: 299 (part or whole).? Chorizopora brongniartii (Audouin) View in CoL : Hincks, 1880: 80.
Material examined. Holotype: MNCN –25.03/3864, 32º38’30.67’’ N, 16º49’47.49’’ W, 18 m, April 2011, on rhodoliths. Paratypes: MNCN –25.03/3860, 3861, NHMUK 2014.1.16.2a: data as in holotype. Other material: NHMUK 1911.10.1.911: Madeira, A.M.R. 1897; NHMUK 1919.6.25.89: Madeira, Norman Coll.; NHMUK 2000.5.30.1: Madeira, Lords of Admiralty; DPO FA, T2005-2A: Departamento de Geología, Universidad de Oviedo, Pajaudina atlantica Logan, 1988 , La Restinga, El Hierro (Canarias) 5– 30 m.
Etymology. From Latin rosarium, rosary, alluding to resemblance of the groups of kenozooids surrounding the autozooids to strings of prayer beads.
Description. Colony encrusting, unilaminar. Autozooid oval to almost rectangular, as wide proximally as distally. Frontal shield flat or slightly convex, without umbo; marked with longitudinal striations and transverse undulations, producing a coarse appearance. Autozooidal orifice surrounded by thin rim, trapezoidal, wider than long, broadest at distal edge, proximal margin straight. Autozooids disjunct, space between them normally filled with small interzooidal kenozooids that sometimes occupy large areas. These oval or irregular, with a rounded central membranous area and tiny opesial opening. Each kenozooid surrounded by 3–6 short communication tubules. Similarly, each autozooid is linked to neighbours (kenozooids or autozooids) by 12–16 such tubules. Each tubule originates from a basal pore-chamber with a uniporous septulum. A single interzooidal avicularium distal to each autozooid, small, elliptical, connected to autozooids and kenozooids by tubules; rostrum triangular, directed distally. Additional avicularia rare, replacing interzooidal kenozoids. Ovicell cleithral, prominent, ooecium longer than wide, formed by flat distal kenozooid ( Fig. 19 View FIGURES 15 – 21. 15 ). Orifice of maternal autozooid dimorphic, clearly larger than in non-fertile ones, transversely D-shaped with straight proximal edge. Ectooecium smooth, with faint radiating striations and sometimes a central medial suture. Proximal edge of ooecium often thickened. All ooecia associated with a small distal avicularium. Ancestrula (observed once) small, oval, with central, smooth frontal surface bearing a T-shaped suture and four frontal spines; its orifice D-shaped, as long as wide, with four articulated oral spines. Ancestrula with six communication tubules, one distal, one proximal and two lateral pairs, connected to one distal, two distolateral, and probably one proximal autozooid, plus to two proximolateral interzooidal avicularia.
SD, Standard deviation; N, number of measurements.
Remarks. Chorizopora rosaria n. sp. is similar to C. brongniartii ( Audouin, 1826) , the only species of the genus reported in the north Atlantic and cited also from Madeira by Hincks (1880) and Norman (1909 as C. annulata ). This species is considered to be cosmopolitan, being absent only in polar waters. However, the existence of so-called cosmopolitan taxa has been not supported for a number of bryozoan species ( Tilbrook 2006; Harmelin et al. 2011; see also Harmelin et al. 2012 for discussion), and it is likely to be the case with C. brongniartii .
Flustra brongniartii Audouin, 1826 was based on a specimen (now lost) from Egypt illustrated by Savigny (1817) (see d’Hondt 2006). The origin of the sample is also unknown, and it may have come from the Mediterranean or from the Red Sea. The original drawings (see Savigny 1817, pl. 10, fig. 6) show a small colony growing on seaweed, with oval, convex, disjunct zooids linked by tubules. The zooids seem to present a large suboral umbo, more developed in ovicellate zooids, projecting over the orifice and concealing it in frontal view. There is no indication of interzooidal avicularia or kenozooids, both considered as characteristic of the genus (see Hayward & Ryland 1999). Finally, despite the small size of the colony, no zooid can be identified as an ancestrula. In all the literature reporting C. brongniartii , as well as material examined by us, this species typically encrusts hard substrata. A distal interzooidal avicularium is always present, as well as kenozooids, although very variable in number, from scattered to abundant, always small and filling spaces between autozooids. The degree of calcification of the zooids also varies, from glistening, translucent, to opaque white, and the texture varies from smooth to coarsely striate. The difference between the original description of F. brongniartii and other species associated with this name, the supposedly wide range of distribution, and the morphological variability, all suggest that more than one species has been described under this name. For instance, the records from Vanuatu ( Tilbrook et al. 2001) and the Solomon Islands ( Tilbrook 2006) probably represent a distinct, undescribed species (see Vieira et al. 2010). As stated by Tilbrook (2006) a more thorough examination of the material assigned to C. brongniartii may illuminate geographical variations or a complex of distinct species. However, the establishment of the specific characters of C. brongniartii , its variability and geographical distribution are beyond the scope of this paper.
We have compared our Madeiran material with specimens and descriptions attributed to C. brongniartii from the Atlantic-Mediterranean region ( Gautier 1962; Arístegui Ruiz 1984; Zabala 1986; Zabala & Maluquer 1986; López de la Cuadra 1991; Hayward & Ryland 1999; Reverter-Gil & Fernández-Pulpeiro 2001; Hayward & McKinney 2002; Souto et al. 2010a, b; Ostrovsky et al. 2013). Madeiran material described here differs in several respects—autozooids are rectangular, flat, lacking an umbo, and have a characteristic coarse surface, while ‘conventional’ C. brongniartii autozooids tend to be more oval, or even subrhomboidal or pear-shaped, convex with a smooth surface, and usually have a suboral or central umbo. In C. rosaria the length of autozooidal orifice is three quarters of the width, the distal avicularium is somewhat separated from the autozooid, and the ooecium has a thickened proximal rim, while in C. brongniartii the length of the orifice is a half of its width, the avicularium is closely apposed to the autozooid, and the ooecium may have a longitudinal frontal ridge. Kenozooids are extremely abundant in Madeiran material, usually surrounding each autozooid, and quite variable in shape and size, while in what is called C. brongniartii these are less frequent, though their number varies among colonies. They are oval and also relatively small. Finally, the zooidal dimensions of our material are much less than in C. brongniartii from the British Isles ( Hayward & Ryland 1999) and from the Adriatic ( Hayward & McKinney 2002) but similar in size to C. brongniartii from the Ría de Ferrol (NW Spain, see Fig. 21 View FIGURES 15 – 21. 15 ).
Taking into account these differences as well as the distance from the supposed type-locality of C. brongniartii (Red Sea or Eastern Mediterranean) we have described our material from Madeira as new. It is possible that C. rosaria has been reported previously as C. brongniartii , but a thorough revision of material will be necessary to determine this. We have revised three samples of C. rosaria from Madeira at the NHMUK (see material examined). One of them originates from the Norman collection and was probably cited by this author ( Norman 1909) as Chorizopora annulata . We have no further information about the record of C. brongniartii made by Hincks (1880). Further studies will be necessary to confirm if C. brongniartii is actually present in Madeira. Chorizopora rosaria n. sp. was also found in the Canaries (see material examined), and was figured in the volume of the Fauna Ibérica series dedicated to Phoronida and Brachiopoda ( Álvarez et al. 2005, fig. 36) together with a specimen of the brachiopod Pajaudina atlantica .
Specimens of C. rosaria from our material encrust maërl rhodoliths, but the colonies at the NHMUK grow on shells, stones and wood; the specimen from the Canaries encrusts a coral.
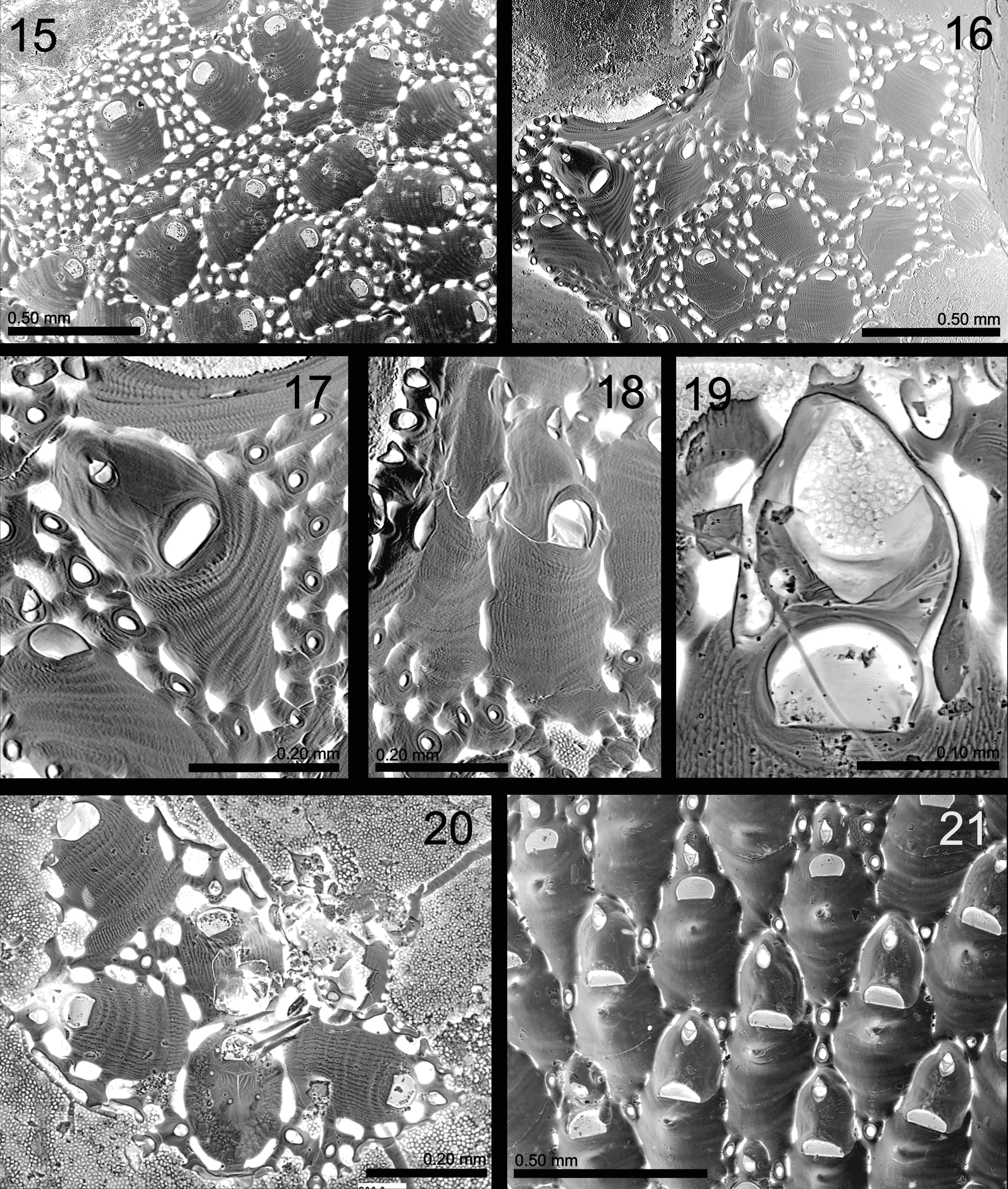
TABLE 3. Measurements (in mm) of Chorizopora rosaria n. sp.
| Mean | SD | Minimum | Maximum | N | |
|---|---|---|---|---|---|
| Autozooid length | 0.340 | 0.0256 | 0.272 | 0.373 | 17 |
| Autozooid width | 0.225 | 0.0225 | 0.184 | 0.266 | 17 |
| Orifice length | 0.056 | 0.0044 | 0.046 | 0.064 | 17 |
| Orifice width | 0.077 | 0.0042 | 0.071 | 0.086 | 17 |
| Avicularium length | 0.043 | 0.0062 | 0.027 | 0.054 | 17 |
| Avicularium width | 0.030 | 0.0048 | 0.024 | 0.042 | 17 |
| Ovicell length | 0.183 | 0.0172 | 0.157 | 0.200 | 10 |
| Ovicell width | 0.173 | 0.0133 | 0.148 | 0.187 | 10 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SubOrder |
Neocheilostomina |
|
Family |
|
|
Genus |
Chorizopora rosaria
| Souto, Javier, Reverter-Gil, Oscar & Ostrovsky, Andrew N. 2014 |
Chorizopora annulata
| Norman 1909: 299 |
| Hincks 1880: 80 |