
Scaphiophryne, INFERRED
|
publication ID |
https://doi.org/10.1111/j.1096-3642.2007.00329.x |
|
persistent identifier |
https://treatment.plazi.org/id/039D87F8-FFBB-FFE5-76B1-FA0F7E7BCE99 |
|
treatment provided by |
Felipe |
|
scientific name |
Scaphiophryne |
| status |
|
RELATIONSHIPS WITHIN SCAPHIOPHRYNE INFERRED View in CoL
FROM LARVAL CHARACTERS
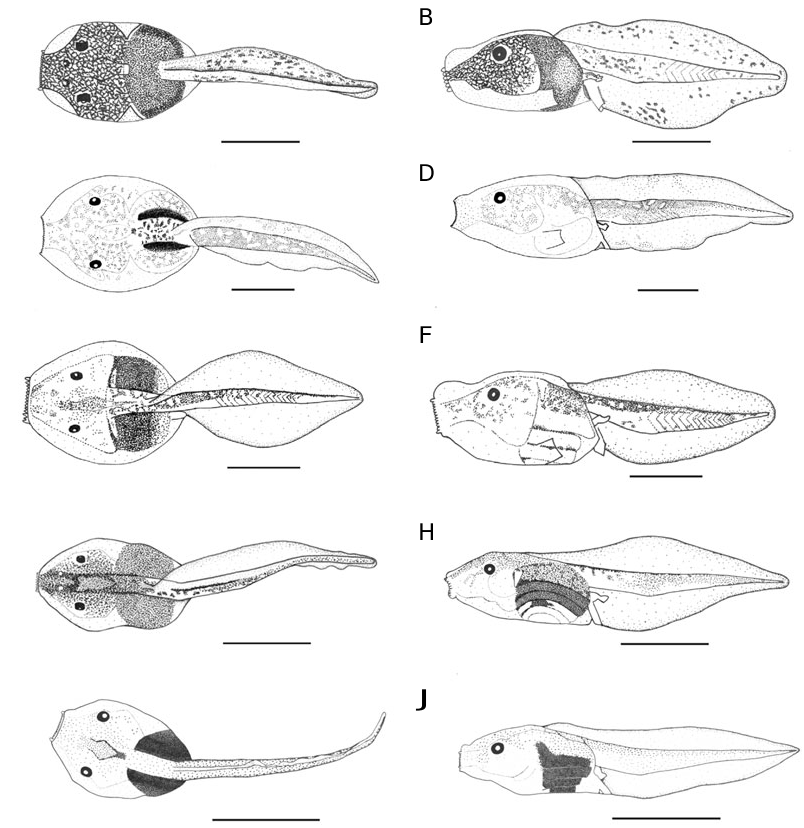
Tadpoles of Scaphiophryne are known to have a unique morphology ( Blommers-Schlösser, 1975; Wassersug, 1984; Haas, 2003; Mercurio & Andreone, 2006), but comparative data on various species were not previously available. Our study provides clear evidence for the existence, within this genus, of two subgroups defined by larval characters of both external and buccopharyngeal morphology, here referred to as the small tadpole group and large tadpole group ( Table 2). Polarizing these characters phylogenetically is difficult as the distinguishing states are not found in the immediate outgroup ( Paradoxophyla ) or are probably of poor phylogenetic value, such as body size and distance of epidermis from inner organs. As Paradox- ophyla tadpoles are small and have no distinct separation of skin from inner organs, the latter two characters can tentatively be used phylogenetically and would indicate the larger size and large distance of skin from inner organs in the large tadpoles group of Scaphiophryne to be synapomorphic. This cavity may possibly represent an enlargement of lymphatic sacs or sinuses (see Viertel & Richter, 1999: 101), and is probably an extreme expression of the state also seen in other microhylids, and in several pipids (in particular in Silurana ; Orton’s tadpole type I). Only speculation is possible regarding the possible adaptive value of this character, but as these tadpoles are very fragile, a protective function is unlikely. Pipid larvae and typical microhylid larvae (Orton’s types I and II) are often floating in midwater, and a function as balancer or float of these additional cavities may be possible. In this case, species of the large tadpoles group of Scaphiophryne may be more strongly adapted to such a floating behaviour, an hypothesis that could be tested by field observations or experimental data.
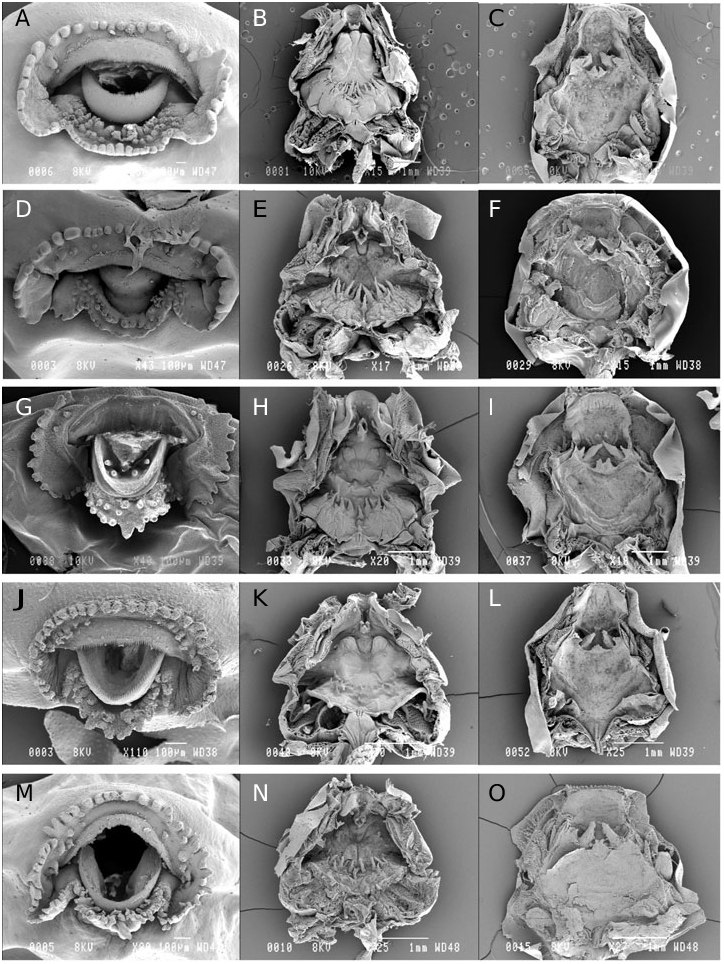
Other larval characters may also be useful to infer relationships among these species, such as the large dorsal bulges sometimes visible on the snout (e.g. S. madagascariensis and S. spinosa in Fig. 1 View Figure 1 ). These bulges are not preservation artefacts and are sometimes also visible in living specimens, but we do not have sufficient data to ascertain their possible diagnostic value. Wassersug (1984) noted that the lower jaw sheath of Scaphiophryne calcarata (under the name Pseudohemisus granulosus ) was serrated and the upper jaw sheath was not serrated. Our data (see Fig. 2 View Figure 2 ) provide clear evidence of very fine serrations on both upper and lower jaw sheaths in all species, including S. calcarata .
The two subgroups can also be identified using adult morphological, bioacoustic and molecular characters. (1) Scaphiophryne brevis and S. calcarata , which are included in the small tadpole group, lack dilated terminal finger discs and have very loud, nonharmonious advertisement calls. (2) Scaphiophryne madagascariensis , S. menabensis and S. spinosa are included in the large tadpole group, and according to the tadpole description of Mercurio & Andreone (2006), the tetraploid species S. gottlebei belongs within this group as well. Indeed, this latter tadpole is of large size (up to 48.2 mm total length in Gosner stage 38), with nares positioned closer to eyes than to tip of the snout, a spiracle lacking an inner wall, and the upper fin not extending onto the body. However the tadpole of S. gottlebei , according to its description in Mercurio & Andreone (2006), may possess an oral disc that differs greatly from those of the other members of the genus. For instance, the emargination is lacking and the submarginal papillae are neatly arranged over the entire oral disc whereas the submarginal papillae are arranged in a similar way in all five tadpoles described herein (i.e. a short row along the lower jaw on the lower labium and a row largely interrupted medially on the upper labium and so composed of only a few submarginal papillae). It needs to be clarified whether these differences reflect biological reality or may just be due to a different perception of the same characters by different researchers.
All four species here included in the large tadpole group are characterized by a very similar advertisement call ( Vences et al., 2003b; Vences, Glaw & Marquez, 2006; Glos et al., 2005), consisting of very rapidly repeated short melodious notes. They also form a very closely related monophyletic group based on molecular data ( Glos et al., 2005), together with the remaining two Scaphiophryne species ( S. boribory and S. marmorata ). Although the calls and larvae of the latter two species are undescribed, we can predict with some confidence that they will be similar in these characters to the other species of the large tadpole group. Within this group, most species are characterized by dilated terminal discs of fingers and toes ( S. gottlebei , S. boribory , S. marmorata , S. menabensis , S. spinosa ) that are largely absent in S. madagascariensis . To some degree, the two subgroups correspond to the subgenera Scaphiophryne Boulenger, 1882 (large tadpole group), with type species Scaphiophryne marmorata Boulenger, 1882 and Pseudohemisus Mocquard, 1895 , with type species Hemisus obscurus Grandidier, 1872 (small tadpole group). We therefore propose to continue to use these subgeneric names but emphasize that the expansion of terminal finger discs is not an unequivocal character to distinguish them. Instead, advertisement calls and especially larval morphology may become the major defining characters once these data are gathered for the two remaining species, S. boribory and S. marmorata .
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.