
Thanatotheristes, Voris & Therrien & Zelenitsky & Brown, 2020
|
publication ID |
https://doi.org/ 10.1016/j.cretres.2020.104388 |
|
publication LSID |
lsid:zoobank.org:pub:26778F25-3135-412D-A203-752CB9C078F1 |
|
DOI |
https://doi.org/10.5281/zenodo.3679538 |
|
persistent identifier |
https://treatment.plazi.org/id/039E87F3-FFE7-FFDA-B92B-DBF56DCEFC17 |
|
treatment provided by |
Plazi |
|
scientific name |
Thanatotheristes |
| status |
gen. nov. |
Thanatotheristes degrootorum gen. et. sp. nov.
ZOOBANK ID: urn:lsid:zoobank.org:act:581C0612-B697-4D01-915C-E3E937D3667A
Etymology. From Thanatos , the Greek god of and embodiment of death, and theristes (Greek), one who reaps or harvests, a reaper. The specific name, degrootorum, is in honor of John and Sandra De Groot of Hays, Alberta, who discovered the holotype specimen and have been supportive of paleontological research in the area.
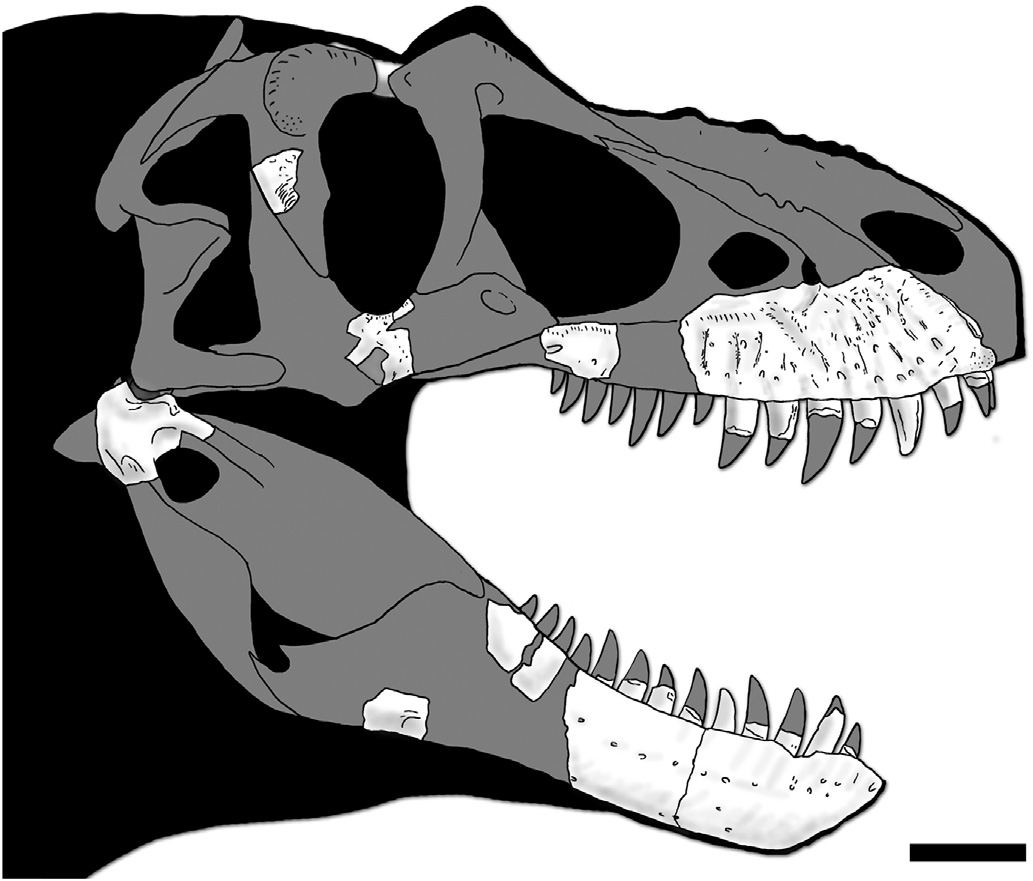
Holotype. TMP 2010.5.7 ( Fig. 2 View Fig ), a partial skull from a large subadult individual (estimated skull length ¼ ~ 800 mm, based on frontal width, metrics from Currie, 2003a) that includes the right maxilla, right jugal, right postorbital, right surangular, right quadrate, right laterosphenoid, left frontal, and both dentaries. Immature bone grain (most apparent within the antorbital fossa of the maxilla) indicates the specimen was not osteologically mature at the time of death ( Carr, 1999), but the well-developed nature of ornamentation on the maxilla suggests the individual was probably sexually mature ( Knell et al., 2013; Hone et al., 2016).
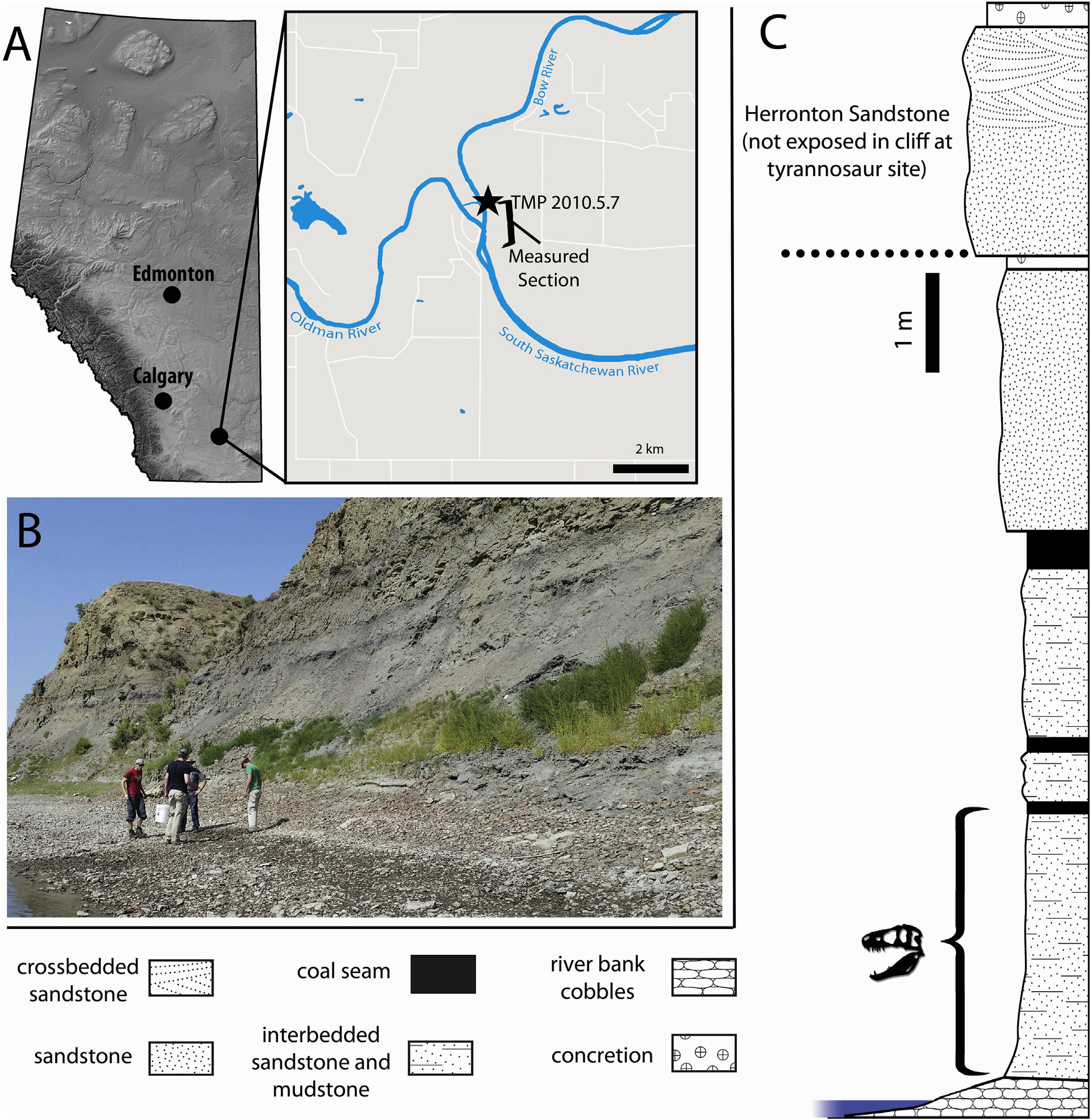
Type locality and horizon. TMP 2010.5.7 was found at the base of a steep cliff on the eastern shore of the Bow River approximately 0.5 km upstream of the confluence with the Oldman River (i.e., The Forks) in Alberta, Canada ( Fig. 3A View Fig ). The cliff is approximately 11 m in height and consists of exposures of the Taber Coal Zone , below the Herronton Sandstone , of the Foremost Formation ( Fig. 3B View Fig ). The exact stratigraphic provenance of the specimen within the Taber Coal Zone is unknown, as the skull was found scattered along the shoreline over a lateral extent of ~ 10 m, but the specimen likely derived from the lower half of the cliff due to a small-scale landslide ( Fig. 3C View Fig ) .
Referred specimen. TMP 2018.16.1, a partial right maxilla from a subadult individual. TMP 2018.16.1 was found along Twelve Mile Coulee, approximately 10 km northeast of Hays, Alberta, stratigraphically high within the Herronton Sandstone of the Foremost Formation.
Diagnosis. Tyrannosaurine tyrannosaurid diagnosed by the following autapomorphies: single row of evenly-spaced dorsoventrally-oriented ridges on the subcutaneous surface of the maxilla ventral and anteroventral to the antorbital fossa; rounded and inflated orbital margin of the jugal; sagittal crest on the frontal extends anterior to the supratemporal ridge as a broad and rounded ridge; lacrimal contact surface on the frontal extends anteromedially at ~60 relative to interfrontal suture; prefrontal with two posteriorly-projecting prongs articulating with the frontal on ventral surface of skull roof (long, medial, primary prong and shorter secondary, lateral prong).
5. Description
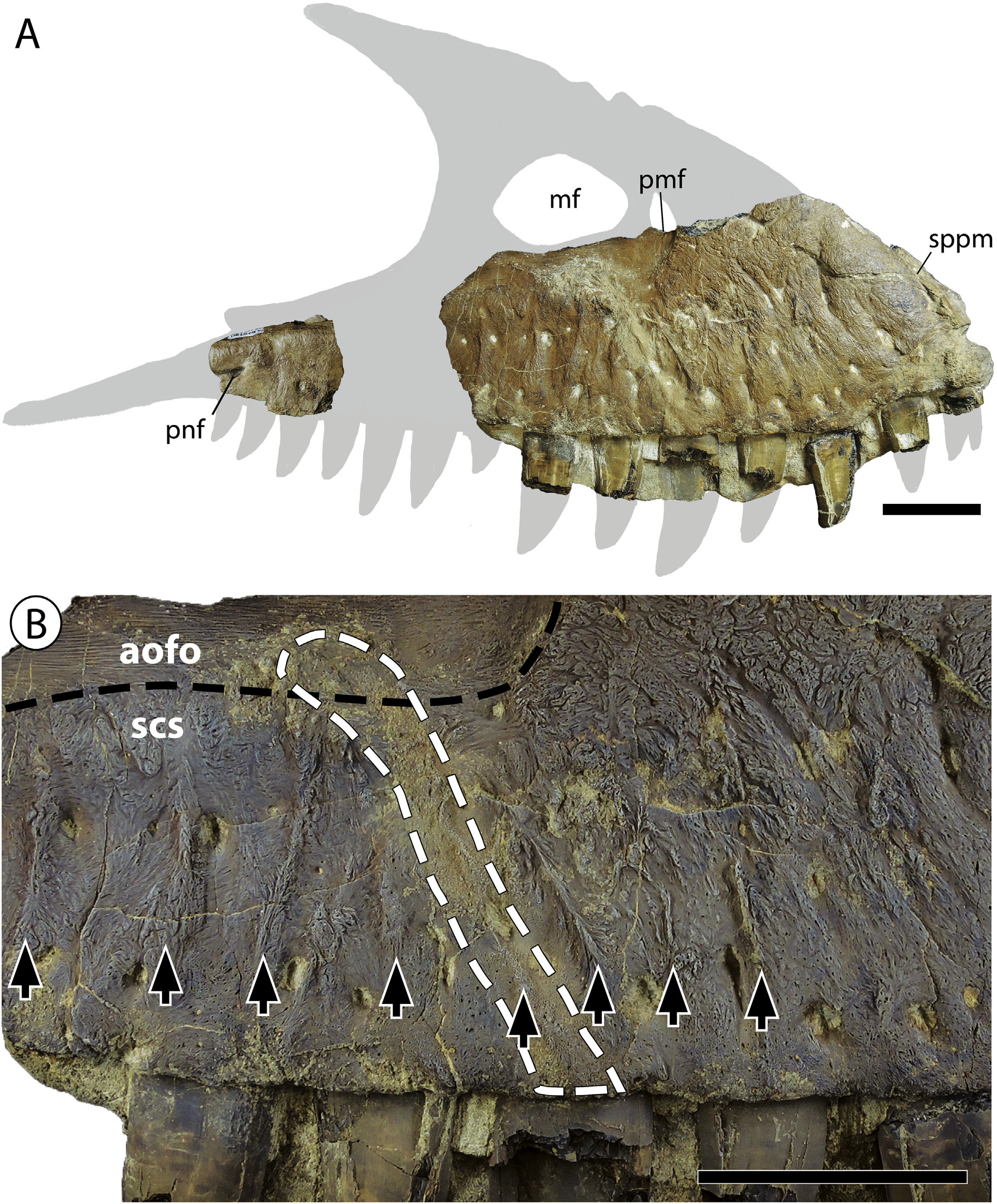
Maxilla -The partial right maxilla of the holotype specimen, TMP 2010.5.7, consists of a complete anterior body and two fragments of the jugal ramus, one of which preserves the three posterior-most alveoli and the groove-like primary posterior foramen of the maxillary nerve and the second preserves a single alveolus ( Fig. 4 View Fig ). The anterior body is deep, consistent with the proportions of other medium to large-sized tyrannosaurines. The anteroventral corner of the bone is slightly downturned and tapers into a shallow angle, as in Daspletosaurus . The referred maxilla (TMP 2018.16.1) consists of the subcutaneous surface ventral to the anterior half of the antorbital fossa ( Fig. 5 View Fig ).
The subcutaneous surface of both specimens is extremely rugose, particularly anteroventral to the antorbital fossa where it is incised by deep anastomosing sulci, as in Daspletosaurus ( Carr et al., 2017) . The subcutaneous texturing differs from that of Daspletosaurus and all other tyrannosaurids in the presence of a single row of prominent and evenly spaced dorsoventrally-oriented ridges located ventral and anteroventral to the antorbital fossa ( Figs. 4 View Fig and 5 View Fig ). In Tyrannosaurus , Tarbosaurus , and Zhuchengtyrannus , the subcutaneous surface is ornamented with at least two rows of unevenly spaced and anastomosing ridges that circumferent shallow oval fossae ( Hone et al., 2011; Carr et al., 2017); such fossae are absent in Thanatotheristes .
The ventral margin of the promaxillary fenestra in TMP 2010.5.7 appears as a wide concavity in the far anterior corner of the antorbital fossa in lateral view ( Fig. 4 View Fig ). The presence of a short base of the preantral strut, however, suggests the exposure of the promaxillary fenestra in lateral view is the result of taphonomic postburial deformation, and that the promaxillary fenestra would have been concealed in lateral view, as in derived tyrannosaurines (e.g., Daspletosaurus , Tarbosaurus , Tyrannosaurus ; Brusatte and Carr, 2016; Carr et al., 2017).
The ventral margin of the maxillary fenestra is broken in TMP 2010.5.7 but its location can be determined based on the thinness of the bone between two thickened areas representing the base of the interfenestral and preantral struts. The ventral margin of the maxillary fenestra is widely separated from the ventral margin of the antorbital fossa, as in albertosaurines, alioramins, Teratophoneus, and juvenile individuals of the genera Daspletosaurus and Tyrannosaurus ( Carr, 1999; Carr et al., 2017). This contrasts with the condition seen in subadult and adult Daspletosaurus individuals, where the ventral margin of the maxillary fenestra is closer to the ventral margin of the antorbital fossa ( Brusatte et al., 2012), and in subadult and adult individuals of more derived tyrannosaurines ( Tyrannosaurus , Tarbosaurus , Zhuchengtyrannus ), where the maxillary fenestra is in contact with the margin of the antorbital fossa ( Carr et al., 2017).
The neurovascular foramina of the maxilla are large in TMP 2010.5.7 and occur in two distinct rows: a dorsal circumfenestral row inset into an elongate fossa and a ventral alveolar row. Anteriorly, the circumfenestral foramina diverge from the linear arrangement with individual foramina becoming randomly dispersed. Several deep and anteroventrally-oriented neurovascular sulci originate from these anterior circumfenestral foramina. The posterior-most foramen of the maxilla exits the subcutaneous surface between the two posterior-most alveoli and continues posteriorly as an elongate neurovascular sulcus, as seen in Lythronax , Daspletosaurus , and Tyrannosaurus . In contrast, the foramen is located posterior to the posterior-most alveolus in albertosaurines.
The jugal ramus of the maxilla in TMP 2010.5.7 is dorsoventrally constricted at the level of the two posterior alveoli, such that the posterior-most maxillary foramen lies near the dorsal margin of the subcutaneous surface ( Fig. 4 View Fig ), as in Daspletosaurus . In mature specimens of all other tyrannosaurids, the jugal ramus gradually tapers posteriorly and the posterior-most maxillary foramen is widely separated from the dorsal margin of the subcutaneous surface.
In total, the anterior body of the maxilla and posterior jugal ramus fragments of TMP 2010.5.7 preserve 12 teeth. However, as these fragments do not articulate with each other and are broadly separated, it is likely that the individual possessed at least 15 maxillary teeth, indicating the animal possessed a long snout ( Fig. 4 View Fig ), as in Daspletosaurus . The base of the tooth crowns are marginally narrower than long (fore-aft base length> crown width). The mesial carinae terminate distal to the base of the crown, which differs from that reported for specimens of Daspletosaurus in which the carinae extend to the base of the crown ( Carr and Williamson, 2000).
The subcutaneous surface of TMP 2010.5.7 is incised by a prominent anteroventral bite mark located immediately ventral to the antorbital fossa (Type 2: sensu Tanke and Currie, 1998). Orientation of the healed bone grain indicates the bite originated at the dorsal extent of the scar and was dragged downward to the alveolar margin at the anterior edge of the sixth alveolus ( Fig. 4 View Fig ). Several possible long-healed Type 1 bite marks (sensu Tanke and Currie, 1998) adorn the subcutaneous surface more anteriorly.
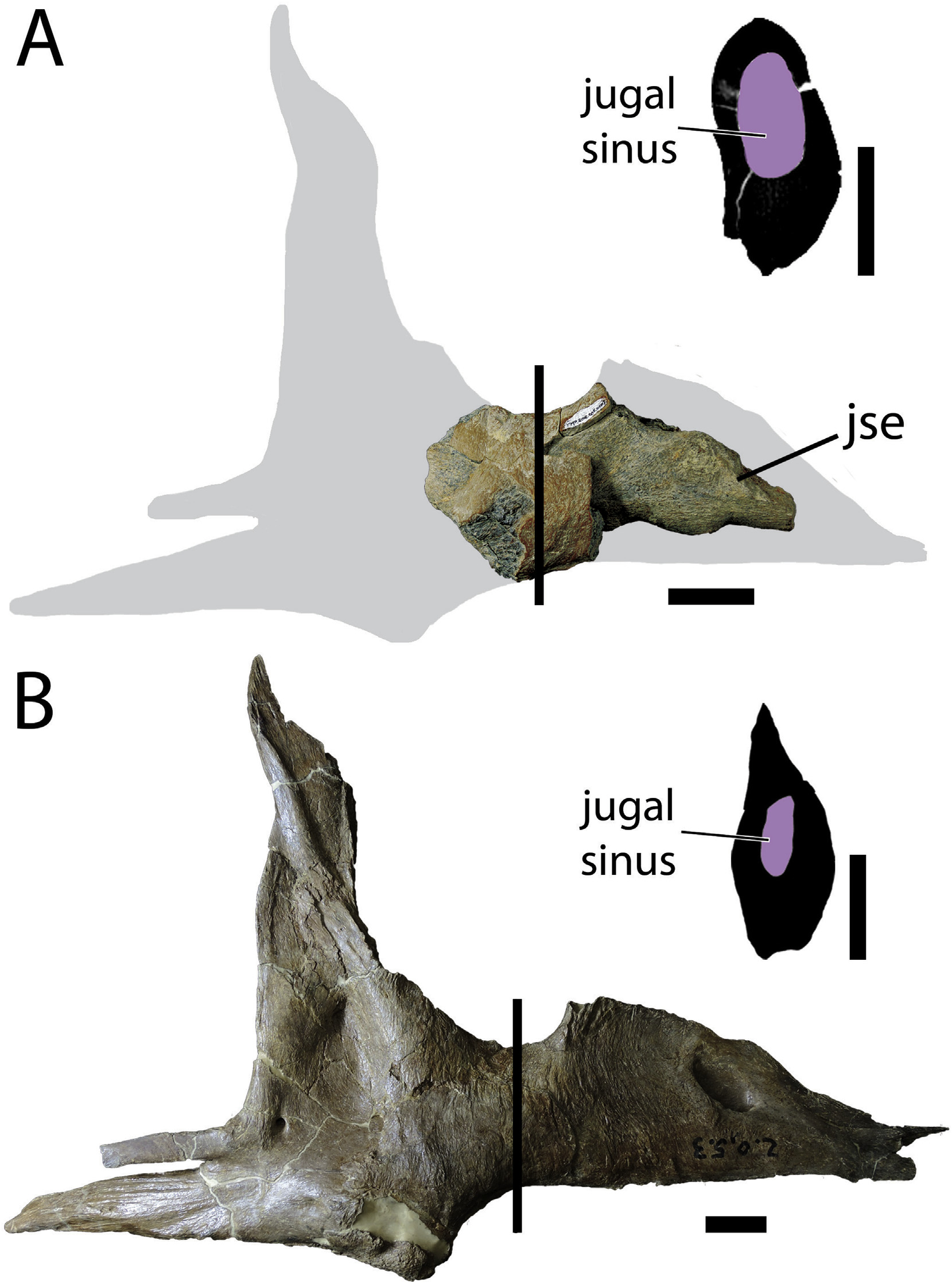
Jugal -The jugal of TMP 2010.5.7 is represented by a fragment of the suborbital region of the bone and a natural endocast of the jugal sinus ( Fig. 6A View Fig and Fig. S3 View Fig ). Despite the fragmentary nature of this bone, the jugal exhibits several features that permit comparison with other tyrannosaurids.
The orbital margin of the jugal is deeply concave and is inclined steeply anterodorsally towards the dorsoventrally-expanded maxillary ramus. The orbital margin is wide and has a rounded edge, resulting in an oval cross-section of the suborbital region ( Fig. 6A View Fig ). This morphology differs from that of most other tyrannosaurids (including Daspletosaurus ), where the orbital margin has a tapered blade-like edge, resulting in a tear drop-shaped crosssection ( Fig. 6B View Fig ). In Lythronax (UMNH VP 20200) and some specimens of Tyrannosaurus rex (TMP 1981.6.1), the jugal suborbital margin is rounded but narrow, differing from the wider margin of Thanatotheristes .
The jugal sinus is greatly inflated in Thanatotheristes , presumably related to the rounder cross-section of the element. The sinus is of consistent depth within the suborbital region, possessing roughly parallel dorsal and ventral margins ( Fig. 6A View Fig ), in contrast to Alioramus ( Gold et al., 2013:fig. 5), Gorgosaurus (TMP 2000.12.11), and Daspletosaurus ( Fig. 6B View Fig ), where the suborbital region of the jugal sinus is dorsoventrally-constricted and hourglass-shaped in lateral view. The lateral surface of the jugal is inflated into a rounded bulge as in Daspletosaurus and most other tyrannosaurids, but unlike in Gorgosaurus and Bistahieversor where it is raised into a shelf-like crest.
Postorbital -The postorbital of TMP 2010.5.7 is represented by a fragment of the ventral ramus coming from the region immediately below the caudal inflection point at the dorsal end of the jugal contact ( Fig. 7 View Fig ). In lateral view, the postorbital is sculptured with a series of parallel and arced ridges and grooves as in other tyrannosaurids. These ridges are separated from the caudal edge of the postorbital by a narrow surface (10 mm) of smooth bone ( Fig. 7A View Fig and Fig. S4 View Fig ).
In medial view, the contact surface for the jugal is braced anteriorly by a robust ridge that extends dorsally to the posterior margin of the ventral ramus ( Fig. 7B View Fig ). Whereas this morphology is seen in some specimens of Tarbosaurus (ZPAL MgD I/4; see Hurum and Sabath, 2003:fig. 8) and Teratophoneus (UMNH VP 16690), the bracing ridge usually terminates at mid-width on the ventral ramus in most tyrannosaurids, or extends to the posterior margin of the ventral ramus as an indistinct, narrow structure in others (e.g. Daspletosaurus , CMN 8506).
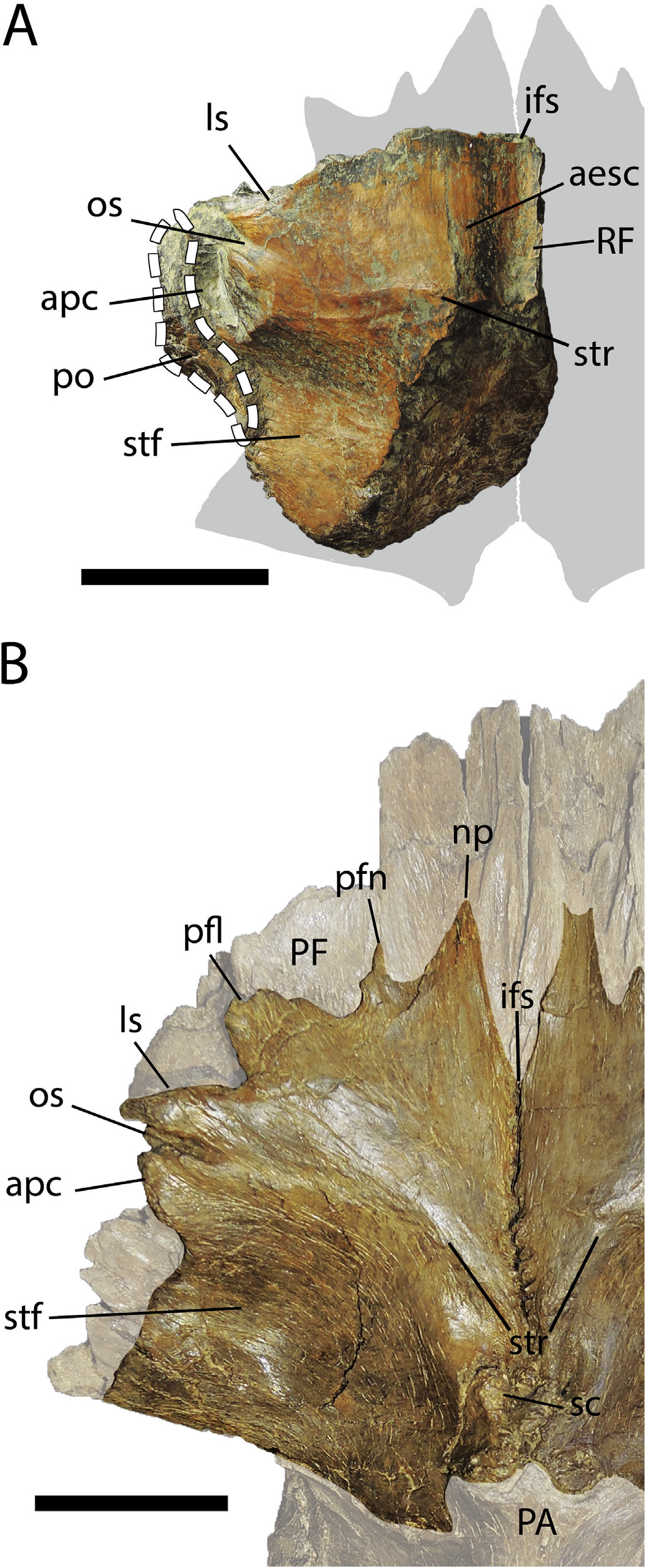
Skull roof -The skull roof of TMP 2010.5.7 consists of a partial left frontal preserved in articulation with a narrow (~ 5 mm wide) fragment of the right frontal, the posteroventral portion of the prefrontal, and small fragments of the postorbital and caudal ramus of the lacrimal ( Fig. 8A View Fig and Fig. S5 View Fig ). The left frontal is missing the prefrontal socket, nasal ramus, most of the sagittal crest, laterosphenoid contact surface, and most of the cerebral fossa on the ventral surface. The contact surface for the postorbital is deep, consistent with that of other derived tyrannosaurines (e.g., Daspletosaurus , Tarbosaurus , and Tyrannosaurus ) but deeper than in other tyrannosaurids (e.g., Albertosaurus , Gorgosaurus, Teratophoneus ).
The dorsal surface of the forehead region (sensu Carr and Williamson, 2004) in TMP 2010.5.7 slopes medially towards the interfrontal suture as in most mature tyrannosaurines, except for Teratophoneus, where it slopes away from the interfrontal suture, as well as in Lythronax and some specimens of Daspletosaurus (e.g., TMP 1980.16.924), where it is flat. The dorsomedial surface is raised into a low, broad, and rounded extension of the sagittal crest located anterior to the supratemporal ridge ( Fig. 8A View Fig and Fig. S5 View Fig ). In all other tyrannosaurids, the sagittal crest does not extend beyond the supratemporal ridge. The supratemporal ridge in TMP 2010.5.7 is sinuous and primarily mediolaterally-oriented, extending from the sagittal crest to the anterior margin of the postorbital contact. Whereas the supratemporal ridge is also sinuous and mediolaterally-oriented in most tyrannosaurids, it is sinuous and anterolaterally-oriented in Daspletosaurus horneri ( Carr et al., 2017) , arcuate and laterally-to anterolaterally-oriented in D. torosus ( Carr et al., 2017) , and straight and mediolaterally-oriented in Tyrannosaurus rex (AMNH FARB 5117).
The dorsal margin of the lacrimal slot on the frontal of TMP 2010.5.7 is oriented at an angle of approximately 60 to the interfrontal suture in dorsal view ( Fig. 8A View Fig ). In contrast, this margin is mediolaterally-oriented in most other tyrannosaurids, except in adult Tyrannosaurus rex (e.g., AMNH 5027, AMNH 5117) where it forms an angle of approximately 70.
In ventral view, the prefrontal-frontal contact is wider and more complex in TMP 2010.5.7 than in all other tyrannosaurids. Whereas the prefrontal typically articulates with the frontal via a single medial prong in tyrannosaurids, it articulates via two prongs, a large medial prong and a shorter, lateral prong, in TMP 2010.5.7 ( Fig. S5 View Fig ). In anterior view, the prefrontal of TMP 2010.5.7 is strongly arced dorsomedially, as in Daspletosaurus where it is related to the presence of a broad, tear drop-shaped prefrontal ( Fig. 8B View Fig ). In contrast, tyrannosaurid taxa in which the arcing of the prefrontal is subtle, extending more dorsally than medially (e.g., albertosaurines, Lythronax, Teratophoneus ), possess a narrow, lens-shaped prefrontal. As such, it can be surmised that TMP 2010.5.7 possessed a broad, tear drop-shaped prefrontal.
Braincase -The braincase of TMP 2010.5.7 is represented by a single fragment of the right laterosphenoid, specifically the antotic region and medial portion of the capitate process, with small fragments of the parietal and right frontal preserved in articulation ( Fig. S6 View Fig ).
The antotic region of TMP 2010.5.7 is broader than in other tyrannosaurids ( Fig. S6 View Fig ). The antotic crest descends ventrally from the capitate process as a prominent, weakly rugose ridge that bifurcates to form a second, more subtle and posteriorly-extending ridge (leading to the otosphenoidal crest in some tyrannosaurids: Bever et al., 2013; Carr et al., 2017). The morphology of the posteriorly-extending ridge contrasts with that of Daspletosaurus , in which it is raised into a wide shelf that extends laterally to the edge of the dorsotemporal fenestra ( Carr et al., 2017).
Quadrate -The right quadrate of TMP 2010.5.7 is represented by a fragmentary mandibular ramus that includes a complete medial condyle and the medial half of the lateral condyle ( Fig. 9 View Fig and Fig. S7 View Fig ). The quadrate pneumatic foramen, located on the ventral anterior face of the quadrate, is ventrally and laterally bounded by a shallow rectangular fossa. The morphology of this fossa is consistent with that of most tyrannosaurids, except in Gorgosaurus where the ventral margin of the fossa inflects dorsally and converges with the lateral wall of the pneumatopore to form a triangular secondary fossa (e.g. TMP 1991.36.500).
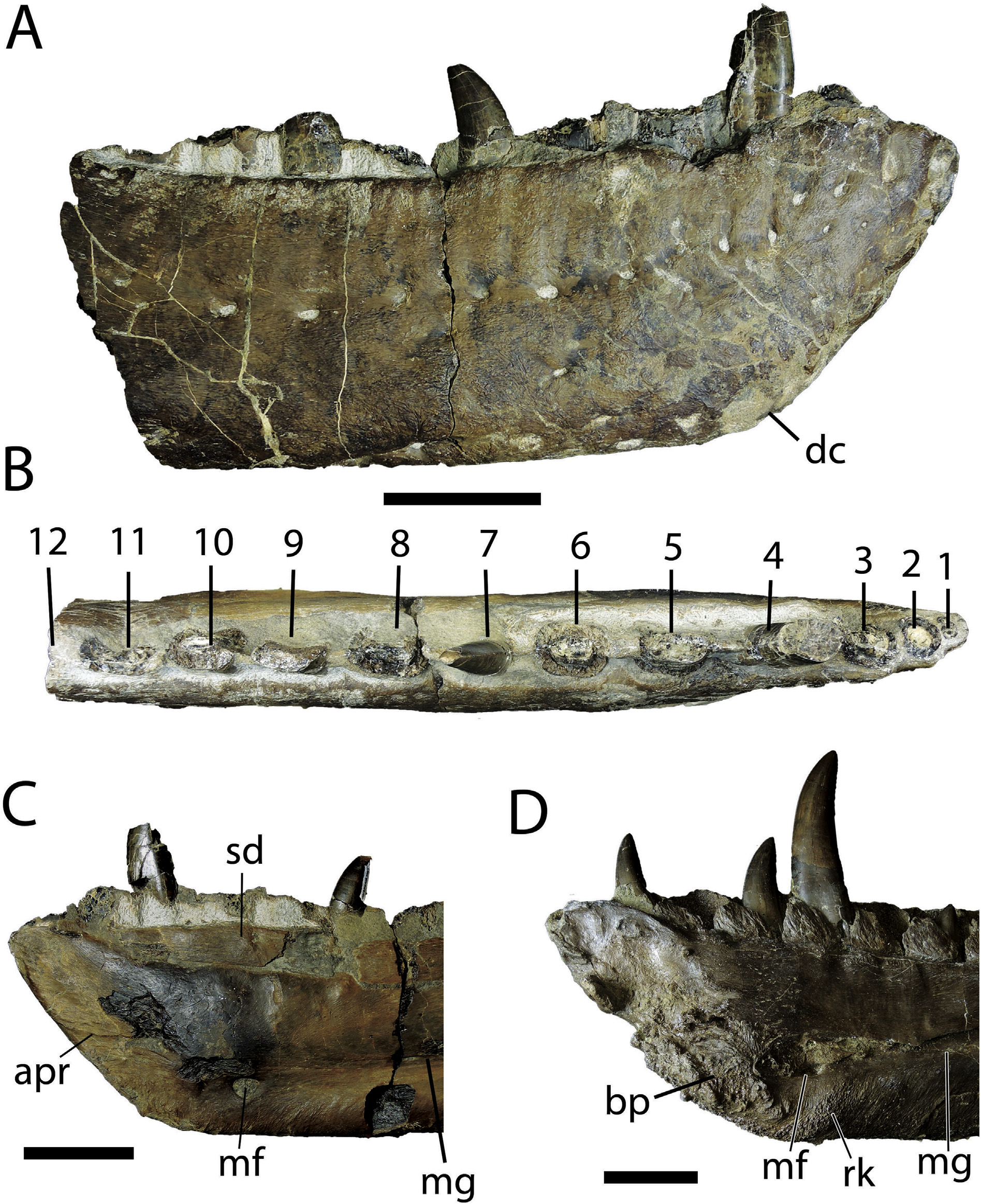
Dentary -The anterior ramus of both dentaries are preserved in TMP 2010.5.7. The left bone preserves the first seven alveoli ( Fig. S8 View Fig ) and the right preserves the first 12 alveoli ( Fig. 10 View Fig ). Two additional fragments of the right dentary are also present, one of the posterior alveolar margin that preserves three additional alveoli (resulting in a total tooth count of 15 teeth) and the other of the splenial joint surface ( Fig. S9 View Fig ). The dorsal margin of both dentaries has been moderately crushed, such that many teeth are broken and fragmentary. Deformation to the main body of the dentaries, however, appears minimal as the Meckelian canal is circular in cross section.
The dentary of TMP 2010.5.7 is nearly of the same depth as that of the largest Daspletosaurus individuals known but much narrower than that of similar-sized or larger individuals ( Fig. 10 View Fig ). The dentary chin (sensu Carr et al., 2017), a subangular to subrounded prominence on the anteroventral margin of the dentary, is located ventral to the third interdental plate as in some Daspletosaurus individuals (ventral to the third alveolus in other Daspletosaurus individuals) and Teratophoneus, but ventral to the fourth alveolus in other tyrannosaurids ( Carr et al., 2017). The dentary chin is more prominent in TMP 2010.5.7 than in either Bistahieversor, Daspletosaurus , or Lythronax , being similar to that observed in albertosaurines and Teratophoneus. The anterior margin of the dentary is straight and inclined anterodorsally at approximately 45, as in albertosaurines and Teratophoneus, but inflects dorsally at the anterior tip of the bone to meet the alveolar margin at an angle of approximately 90 ( Fig.10 View Fig ), as in derived tyrannosaurines ( Carr et al., 2017). Despite its narrowness, the overall dentary morphology is consistent with that of derived tyrannosaurines.
The subcutaneous surface of the dentary is smooth posteriorly and becomes increasingly rugose anteroventrally. The anterior alveolar margin is excavated by shallow, dorsoventrallyoriented neurovascular sulci that originate from the alveolar row of neurovascular foramina. Whereas similar subcutaneous texturing is observed in nearly all tyrannosaurids, the neurovascular sulci of the anterior alveolar margin tend to be deeper in larger, derived tyrannosaurines, such as Daspletosaurus and Tyrannosaurus .
In TMP 2010.5.7, the medial surface of the dentary is incised by a deep Meckelian groove that shallows anteriorly into a sulcus, a characteristic previously interpreted as an indicator of immaturity ( Carr et al., 2017). The ventral bar is swollen and cylindrical posteriorly and flattens anteriorly towards the large (10 mm in diameter), circular Meckelian foramen. The area ventral to the Meckelian foramen is flat and weakly rugose in TMP 2010.5.7 ( Fig. 10C View Fig ), as in albertosaurines, Lythronax , and Teratophoneus, but in contrast to other derived tyrannosaurines ( Daspletosaurus , Tarbosaurus , Tyrannosaurus , Zhuchengtyrannus ) where it is raised into a swollen and rugose knob ( Fig. 10D View Fig ; Carr et al., 2017). The symphyseal surface of TMP 2010.5.7 is rugose ( Fig. 10C View Fig ), bearing prominent anteroposteriorly-oriented ridges as in Tarbosaurus and Tyrannosaurus , but lacks the large and deeply interlocking bony projections observed in subadult and adult individuals of Daspletosaurus ( Fig. 10D View Fig ). Although development of the rugose knob and the symphyseal rugosities may be related to ontogeny ( Carr et al., 2017), the fact that they are present in Daspletosaurus individuals of similar size to TMP 2010.5.7 suggests that their absence in the latter is not due to ontogeny in this specimen.
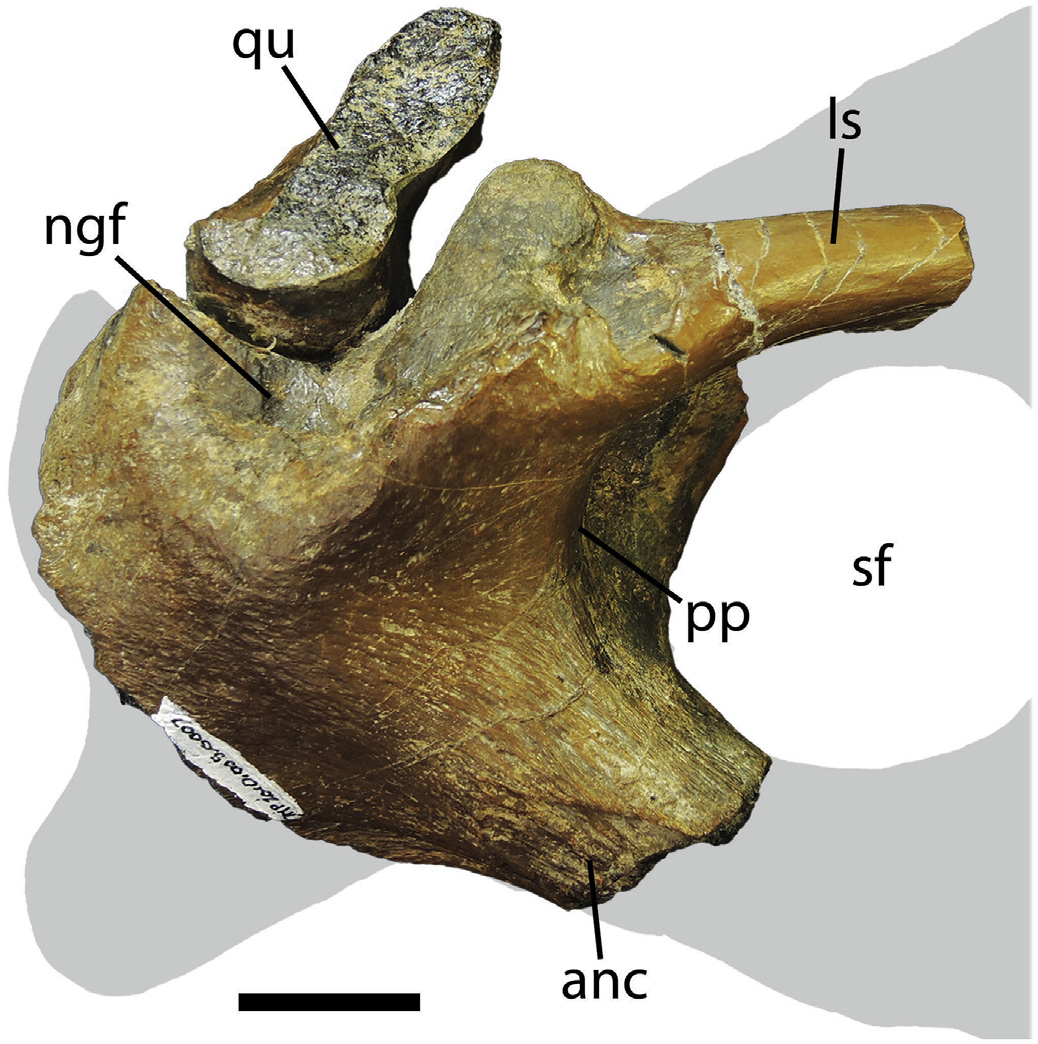
Surangular -Only the posterior-most portion of the right surangular is present in TMP 2010.5.7, which includes the articular ramus, the posterior third of the surangular shelf, and the posterior margin of the surangular foramen ( Fig. 9 View Fig ). A largely undiagnostic portion of the right articular is preserved in articulation with the surangular.
The surangular shelf (sensu Currie, 2003b) is a deep and robust flange located anterior to the quadrate-surangular articulation. In TMP 2010.5.7, the surangular shelf projects dorsolaterally, such that its ventral surface is visible in lateral view ( Fig. 9 View Fig ). In contrast, the surangular shelf projects laterally in albertosaurines and alioramins, and ventrolaterally in derived tyrannosaurines, where it overhangs the dorsal margin of the surangular foramen ( Brusatte et al., 2011). The adductor fossa, located on the dorsal surface of the surangular shelf, is medially bounded by the coronoid process. In TMP 2010.5.7, the coronoid process extends to the posteromedial margin of the adductor fossa and converges on the triangular process anterodorsal to the glenoid fossa ( Fig. S7 View Fig ), as seen in Lythronax (UMNH VP 20200). In all other tyrannosaurids, the coronoid process terminates anterior to the posterior margin of adductor fossa.
A pneumatic pocket occurs posterior to the surangular foramen in all mature tyrannosaurines and most albertosaurine specimens. In TMP 2010.5.7, this pneumatic pocket is enclosed posterolaterally by a thick flange ( Fig. 9 View Fig and Fig. S8 View Fig ). In other derived tyrannosaurines (including Daspletosaurus ), the pneumatic pocket is enclosed posterolaterally and ventrolaterally by this same flange, and dorsolaterally by the posterior edge of the lateral surangular shelf.
The nonglenoid fossa is a crescentic depression located ventrolateral to the glenoid fossa ( Fig. 9 View Fig ; Brusatte et al., 2012). In TMP 2010.5.7, the anteroventral margin of the nonglenoid fossa is bounded by a prominent and rugose crest. This crest is significantly lower in all other tyrannosaurids.


| TMP |
Transvaal Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |