
Liopeltis pallidonuchalis, ı Tan Van Nguyen & Vogel, 2019
|
publication ID |
https://doi.org/ 10.1080/00222933.2019.1656784 |
|
DOI |
https://doi.org/10.5281/zenodo.3671282 |
|
persistent identifier |
https://treatment.plazi.org/id/03A087B3-FFAD-FFEB-009D-2669FF6B5FF7 |
|
treatment provided by |
Valdenar |
|
scientific name |
Liopeltis pallidonuchalis |
| status |
sp. nov. |
Liopeltis pallidonuchalis sp. nov.
( Figures 1 – 5 View Figure 1 View Figure 2 View Figure 3 View Figure 4 View Figure 5 ; Tables 1 – 2 View Table 1 View Table 2 )
Chresonymy
Liopeltis frenatus (partim) – Smith (1943): 183; Ziegler et al. (2007): 10 – 11ı Figure 11 (?); Ziegler and Hoang (2009): 115.
Holotype. ZMMU R-15682ı adult male (hemipenis everted) from forest trail in the evergreen montane tropical forest in Kon Chu Rang N. R. (14.5034° Nı 108.5383° Eı at elevation of 1010 m asl)ı Gia Lai Provinceı central Vietnam ( Figures 2 – 4 View Figure 2 View Figure 3 View Figure 4 )ı collected by Nikolay A. Poyarkov on 29 May 2016.
Paratypes. DTU 307 (adult female) roadkill specimen from the road in the evergreen montane tropical forest in Bach Ma N. P. (16.2035° Nı 107.8540° Eı at an elevation of 950 m asl) ı Thua Thien – Hue Provinceı central Vietnam ( Figure 5 View Figure 5 )ı collected by The Anh Nguyen and Phuong Nhu Hoang on 18 June 2018; ZFMK 83105 About ZFMK (adult female) from Bach Ma N .P. (no elevation data available) ı Thua Thien – Hue Provinceı central Vietnamı collected by Quang Xuan Hoang and students before August 1998 .
Referred materials
VNUH 9.7. ’ 06 – 1 (adult male) roadkill found on the road during the dry season in the U Bo regionı Phong Nha – Ke Bang N.P. ı Quang Binh Provinceı central Vietnam (at an elevation of 600 m asl)ı collected by Astrid Heidrich on 9 July 2006 .
Diagnosis. The new species can be separated from its congeners by the following combination of morphological characters: (1) one single (or missing) loreal; (2) one single nasal shield; (3) dorsal scales in 15-15-15 rowsı all smooth; (4) 1 preocuları 2 postoculars and 1 + 2 temporals; (5) 7 supralabialsı of which 3d and 4th in contact with the eye; (6) 8 infralabials; (7) nasal contacting internasal; (8) prefrontal touching or separated from supralabials; (9) ventral scales 126 – 138; (10) subcaudal scales 67 – 73ı paired; (11) relative tail length about 0.274 – 0.301; (12) an uniform bronze body colouration; (13) a thin postocular stripe extending from eye to end of the neck becoming indistinct posteriorly. The new species can be distinguished from all other congeners by its low subcaudal scales countı and by its characteristic colouration. Detailed comparisons with other species of the genus Liopeltis appear below.
Description of holotype. Body slender and elongate ( Figure 2 View Figure 2 (a))ı somewhat laterally compressed; tail thinı pointed; head smallı moderately distinct from the neck (mostly in dorsal view); snout blunt; nostrils rather largeı in dorsal-lateral positionı round in shape; eye rather largeı pupil circular.
Body size. HL: 19.8 mm; SVL: 297 mm; TaL: 112 mm; TL: 409 mm; ratio TaL/TL: 0.274.
Body scalation. Dorsal scale rows 15-15-15ı all smooth; scales on dorsum and flanks somewhat rhomboid in shape; scales of the vertebral row not enlarged; no apical pit detected; 133 ventrals (+2 preventrals); 67 subcaudalsı all paired; anal plate divided.
Head scalation. Rostral heptagonalı wider than highı slightly visible from above ( Figure 2 View Figure 2 (c)); nasal singleı elongated; nasal surrounded by the first two supralabialsı rostralı internasalı and prefrontal; internasals twoı curvedı wider than longerı in contact with rostral anteriorlyı nasalı and prefrontal; prefrontals twoı prefrontal length comprising less than half of frontal length; prefrontals in contact with internasalsı nasalsı preocularsı and frontal; frontal pentagonalı ca. 1.11 times wider than longı tapering posteriorlyı subequal to the distance from tip of snout to frontal; parietals longer than wideı in contact approximately the length of frontal ( Figure 2 View Figure 2 (c))ı ca. 1.66 times longer than frontal; 1/1 supraocuları distinctly wider than highı in contact with prefrontal; loreal missing; 1/1 preocuları largeı pentagonalı higher than wideı in broad contact with prefrontal; subocular absent; 2/2 small postocularsı upper postocular larger than lower; 7/7 supralabialsı third and fourth entering orbitı sixth largest; 1 + 2/1 + 2 temporalsı anterior one very long and narrow ( Figure 2 View Figure 2 (b))ı in broad contact with supralabials 5 – 6 and parietalsı upper posterior temporal smaller than lower posteriorı rhomboid; infralabials 8/8 ( Figure 2 View Figure 2 (d))ı first pair in contact behind small mentalı 1 – 4/ 1 – 4 in contact with anterior chin shieldsı fifth infralabial largestı apex directed posteriorly; posterior chin shields longer than anterior onesı separated from each other by a pair of small scales ( Figure 2 View Figure 2 (d)).
Dentition. Maxillary teeth smallı equalı 20 on left and 22 on the right side (counted directly prior to holotype preservation).
Hemipenis morphology. In everted positionı the organ is singleı not forkedı stoutı and reaches SC 5. Hemipenis is somewhat dumbbell-shapedı its proximal part covered with numerous tooth-shaped proximally oriented hooks ( Figure 3 View Figure 3 )ı located in 3 – 5 rows; middle and distal parts of the organ covered with over 30 transverse skinfolds each bearing numerous papillae ending with small horny spinesı which increases in size distally; hemipenis tip with rare spines; sulcus spermaticus distinctı prominent ( Figure 3 View Figure 3 (b)).
Colouration (in life). In life body and flanks uniform bronze-brown ( Figures 2 View Figure 2 and 4 View Figure 4 )ı ventrally gradually getting lighter to light-beige background colour of lower flanks; head dorsally somewhat darker bronze-brownı snout beigeı supralabials and ventral surfaces of head immaculate cream-white; pupil blackı iris bicoloured: dorsally goldenı laterally and ventrally copper red-brown ( Figure 2 View Figure 2 (b)); a narrow black postocular stripe of the same width as lower postocular running from the posterior corner of eye along lower postocuları lower margins of primary and lower secondary temporalsı dorsal margins of supralabials 6 – 7; postocular stripe gently curves dorsally at the level of head basis and becomes less distinct in nuchal area; continues as a pair of short dark grey markings with indistinct borders ca. 5 dorsal scales in length in the neck region ( Figure 4 View Figure 4 ). Rare black spots of irregular shape located in the area of mouth angle ( Figure 2 View Figure 2 (b))ı posteriorly spots getting elongated and form a narrow-interrupted dark line running along the border between ventrals and dorsal scales for the anterior one-third of body length ( Figures 2 View Figure 2 (a) and 4); a shorter and less distinct dark line running from the mouth angle along the border between the second and third rows of dorsal scales for approximately the distance of head length and quickly becoming indistinct ( Figure 2 View Figure 2 (a)).
Colouration (in preservative). After 3 years in preservative the general colouration pattern did not change; orange tint in the colouration of dorsumı head and eye faded becoming greyish-brown; other features of colouration remain unchanged.
Variation. Variation in measurements and scalation of the type series is presented in Table 2 View Table 2 . The holotype from Gia Lai Province lacks loreal and prefrontal touches second supralabial ( Figure 2 View Figure 2 (b))ı while in two paratypes from Thua Thien – Hue Province loreal is present and prefrontals are not in contact with supralabials ( Table 2 View Table 2 ) . The paratype DTU 307 has a single small scale separating posterior chin shieldsı while the holotype and the paratype ZFMK 83105 About ZFMK have two small scales anterior to ventrals . Colouration of the paratype DTU 307 is very similar to the colouration oft he holotype described above; but a series of 19 small dark-brown paravertebral blotches with indistinct borders is running from the neck region along with the anterior one-third of body length ( Figure 5 View Figure 5 ) .
A single road-kill specimenı originally identified as L. frenata ı was reported from the lowland karst forest of Phong Nha – Ke Bang N.P. in Quang Binh Province in central Vietnam (elevation 150 m asl) by Ziegler et al. (2007) . In generalı morphological data for this specimen agrees well with the diagnosis of Liopeltis pallidonuchalis sp . nov. with exception of the following differences (see Table 2 View Table 2 ): comparatively longer tail ( Rel TL 0.301 vs. 0.274 – 0.288 in the type series of the new species)ı four small scales between chin shields anterior to ventrals (vs. 1 – 2 scales in the type series of the new species); seven infralabials (vs. eight infralabials in the type series of the new species) GoogleMaps . Taking in the account morphological (see above) and ecological (lowland karst forest vs. montane evergreen forest) differences between these two populationsı we tentatively assign the Quang Binh population to Liopeltis pallidonuchalis sp . nov. as referred materialsı pending examination of additional specimens and molecular data.
Etymology. The specific name ‘pallidonuchalis’ is a Latinised adjective in the nominative singular (feminine gender)ı derived from Latin ‘pallidus’ for ‘ pale ’ and Medieval Latin ‘nucha’ ı derived from Arabic ‘nuka’ for ‘ nape ’ ı ‘ dorsal surface of neck ’ ı referring to the pale postocular marking of the new species disappearing in nuchal area.
We suggest the following common names: Pale-necked Ringneck (English)ı Rắn đai gáy nhạt màu (Vietnamese)ı Blednyi Gladkiy Kamyshovyi Uzh (Russian)ı Blasse Halsbandnatter (German).
Distribution. The distribution of the new species is shown in Figure 1 View Figure 1 . Liopeltis pallidonuchalis sp. nov. is presently known from montane evergreen tropical forests of Kon Tum – Gia Lai Plateau of Truong Son (also known as Annamite) Mountains in Central Vietnam. It is relaibly recorded from Kon Chu Rang N.R. in Gia Lai Province (our data)ı from Bach Ma N.P. in Thua Thien – Hue Province (our data; Ziegler and Hoang 2009) and from Ba Na – Nui Chua N.R. in Da Nang City (historical record from ‘ Tourane ’ ı Smith 1943); where it was found at elevations of 950 – 1010 m asl. The new species probably also occurs in karst forests of Phong Nha – Ke Bang N.P.ı Quang Binh Province ( Ziegler et al. 2007; see Variation) where it occurs at elevations 150 m aslı however this locality requires further verification. In particuları records from eastern Laos are anticipated.
Conservation status. Liopeltis pallidonuchalis sp. nov. is to date reliably recorded only from two national parks and one nature reserve in central Vietnam and is known only from four specimens. This is a highly secretive snake which appears to be a strict forest specialistı associated with primary undisturbed montane forests. It appears that the new species may be affected by growing anthropogenic pressure and forest destructionı as observed in different areas of central Vietnam. Given the available informationı we suggest Liopeltis pallidonuchalis sp. nov. be considered as a Vulnerable (VU) species following IUCN ’ s Red List categories ( IUCN Standards and Petitions Subcommittee 2016).
Biology. Our knowledge of the biology of Liopeltis pallidonuchalis sp. nov. is scarce; the species appears to be closely associated with montane evergreen forests covering the Kon Tum – Gia Lai Plateau of Truong Son Mountains where it was recorded at elevations of 950 – 1010 m asl. It also possibly occurs in lowland karst tropical forests of Quang Binh karst area (see Variation) where the species was recorded from 150 m asl. Animals were recorded only in patches of primary undisturbed forest with complete multi-layered canopy and heavy undergrowthı suggesting the new species is a strict forest-dwelling specialist. At the type locality in Kon Chu Rang N.R. (Gia Lai Province)ı the forest where the new species was recorded is dominated by large trees of the families Podocarpaceae ( Dacrydium elatum , Dacrycarpus imbricatus )ı Magnoliaceaeı Burseraceae ( Canarium sp.)ı Myrtaceae ( Syzygium sp.)ı Hamamelidaceae (Simingtonia sp.)ı Lauraceae ( Litsea sp.)ı Rhodoliaceae (Rhodolia sp.)ı Fagaceaeı Sterculiaceae ( Scaphium sp.) ( Figure 6 View Figure 6 ).
As with other species of the genusı Liopeltis pallidonuchalis sp. nov. was found to be a very secretive speciesı with terrestrial and nocturnal activity; the only specimen that we received alive was recorded at dusk around 19:30 h at an air temperature of 20 – 22°C and 100% humidity; three other known specimens are roadkills. The holotypes stomach contained remains of a spiderı which we tentatively identified as a member of the family Lycosidae Gen. sp. The adult female ZFMK 83105 described by Ziegler and Hoang (2009) contained four eggs of 25 – 28 mm in length and 7 – 9 mm in width. Nothing else is known about the biology of the new species.
Other species of snakes recorded syntopically with the new species at the type locality in Kon Chu Rang N.R. (Gia Lai Province) included Rhabdophis nigrocinctus (Blyth) ı Sinonatrix percarinata (Boulenger) ı Ovophis monticola (Günther) ı Pareas hamptoni (Boulenger) ı Hebius leucomystax (Davidı Bainı Nguyenı Orlovı Vogelı Vu & Ziegler) and Trimeresurus vogeli Davidı Vidal & Pauwels. In Bach Ma N.P. (Thua Thien – Hue Province) the new species was recorded in sympatry with Calamaria concolor Orlovı Nguyenı Nguyenı Ananjeva & Hoı C. pavimentata Dumérilı Bibron & Dumérilı Dendrelaphis ngansonensis (Bourret) ı Lycodon fasciatus (Anderson) ı Oligodon catenatus (Blyth) ı Ptyas carinata (Günther) ı P. korros (Schlegel) ı P. nigromarginata (Blyth) ı Hebius leucomystax , Pseudoxenodon macrops (Blyth) ı Sibynophis cf. collaris (Gray) ı Bungarus slowinskii Kuchı Kizirianı Nguyenı Lawsonı Donnelly & Mebsı Ophiophagus hannah (Cantor) ı Pareas hamptoni , Ovophis monticola , Protobothrops cornutus (Smith) ı and Trimeresurus vogeli .
Comparisons. Comparative morphological data for the new species and currently recognised members of the genera Liopeltis and Gongylosoma is presented in Table 1 View Table 1 . We decided to include the new species into the genus Liopeltis rather than Gongylosoma for the following reasons: the number of dorsal scalesı and the prefrontal touching the upper labialsı both characters are unknown in the genus Gongylosoma . According to Leviton (1964) the head shape of the new species resembles the genus Gongylosoma . Howeverı Leviton did only compare the two species G. baliodeirum ( Boieı 1827) and L. tricolor . Several other Liopeltis species do rather have the ‘ shortı deep and convex ’ head profile than the ‘ long shallowed and flattened ’ shape which Leviton defined for the genus Liopeltis . Presentlyı based entirely on morphological reasons we are not convinced about the necessity of separation of these two genera.
Morphologically Liopeltis pallidonuchalis sp. nov. is most closely resembling L. calamaria and L. frenata . The main characters which separate it from L. calamaria is the colouration; the fact that the nasals are fused with the internasals in L. calamaria and the prefrontal is widely separated from the prefrontal by the postnasal. Beside that there is a huge distribution gap between the Indianı Sri Lankan and Nepalese localities of L . calamaria and the type locality of Liopeltis pallidonuchalis sp. nov. ı which makes a conspecifity very unlikely.
The main differences of the new species from L. frenata are the lower number of ventralsı subcaudals and the colouration as well. For photos of the type specimen of Cyclophis frenatus Güntherı 1858 (BMNH 1946.1.1.72) see Figure 7 View Figure 7 . Liopeltis pallidonuchalis sp. nov. ı differs from L. frenata by a significantly lower number of ventrals (126 – 138 vs. 140 – 172). The difference in the number of subcaudals seems to be slimı but in L. frenata has a large sexual dimorphism in the number of subcaudals. So the lower part of the range of the subcaudals in L. frenata corresponds to female specimens and the lowest number is given by Boulenger (1890) as 87 – 96 in both sexesı by Smith (1943) and Das (2010) as 70 – 105 in both sexesı by Deuve (1970) as 87 – 103 in both sexesı by Orlov et al. (2003) as 84 in the maleı by Yang and Rao (2008) as 93 in the femaleı by Pham et al. (2014) as 100 in the maleı 94 in the female. We counted 99 in a single male of the holotype of L. frenata ı which is much higher than the 67 – 69 in the males and 71 – 73 in the females of Liopeltis pallidonuchalis sp. nov. In Liopeltis pallidonuchalis sp. nov. the postocular stripe is thin whereas it is very thick in L. frenata (see Figures 7 – 8 View Figure 7 View Figure 8 ). Besides the postocular stripeı in L. frenata 3 – 4 narrower lateral dark stripes are presentı which run from the neck region posteriorly along the anterior half of the body ( Figures 7 View Figure 7 and 8 View Figure 8 ); such dark markings are absent in the new species. Finallyı molecular data suggest that there is a deep divergence between cyt b gene sequences of L. frenata sensu stricto from northern Myanmar and Liopeltis pallidonuchalis sp. nov. (p = 15.3 – 15.6%)ı which is exceeding the species-level of divergence in colubrid snakes.
Liopeltis pallidonuchalis sp. nov. differs from L. philippina by having regularly one loreal (vs. missing); 7 supralabialsı 3rd and 4th in contact with the eye (vs. 8 supralabialsı fourth and fifth in contact with the eye); slight lower number of ventral scales (126 – 138 vs. 139 – 150)ı lower number of subcaudals (67 – 73 vs. 110 – 119)ı dorsal scales in 15-15-15 rows (vs. 15-15-13 rows)ı postocular stripe is thin (vs. four longitudinal brown stripes begin on neck and continue along body). Liopeltis pallidonuchalis sp. nov. differs from L. rappi by having one nasal (vs. two nasals)ı temporals 1 + 2 (vs. 1 + 1)ı higher number of supralabials (7 vs. 6)ı lower number of ventral scales (126 – 138 vs. 176 – 195)ı a thin postocular stripe (vs. none); Liopeltis pallidonuchalis sp. nov. differs from L. stoliczkae by having 7 supralabialsı 3rd and 4th in contact with the eye (vs. 8 supralabialsı fourth and fifth in contact with the eye); lower number of ventral scales (126 – 138 vs. 148 – 154); lower number of subcaudals (67 – 73 vs. 116 – 134); dorsal scales in 15-15-15 rows (vs. 15- 15-13 rows); Liopeltis pallidonuchalis sp. nov. differs from L. tricolor by having regularly one loreal (vs. missing); slight lower number of ventral scales (126 – 138 vs. 140 – 187)ı lower number of subcaudals (67 – 73 vs. 103 – 137)ı dorsal scales in 15-15-15 rows (vs. 15- 15-13 rows)ı postocular stripe is thin (vs. very thick).
Liopeltis pallidonuchalis sp. nov. differs from members of the genus Gongylosoma as follows: G. baliodeirum by having one nasal (vs. two)ı dorsal scales in 15-15-15 rows (vs. 13-13-13 rows)ı 7 supralabialsı third and fourth in contact with the eye (vs. 6 – 7 supralabialsı fourth and fifth in contact with the eye)ı a thin postocularstripe (vs. none); G. longicaudum ( Petersı 1871) by having dorsal scales in 15-15-15 rows (vs. 13-13-13)ı lower number of subcaudals (67 – 73 vs. 91 – 120)ı postocular streak thin and dark (vs. light)ı no stripes at least anteriorly (vs. stripes present); G. mukutense Grismerı Das & Leongı 2003 by having one nasal (vs. two nasals)ı dorsal scales in 15-15-15 rows (vs. 13- 13-13 rows)ı temporals 1 + 2 (vs. 1 + 1)ı lower number of subcaudals (67 – 73 vs. 99)ı postocular streak thin and dark (vs. light)ı no stripes at least anteriorly (vs. stripes present); G. nicobariense ( Stoliczkaı 1870) one nasal (vs. two nasals)ı dorsal scales in 15-15-15 rows (vs. 17 – 17-17 rows)ı regularly one loreal (vs. missing)ı temporals 1 + 2 (vs. 2 + 2)ı lower number of ventrals (126 – 138 vs. 192)ı lower number of subcaudals (67 – 73 vs. 84)ı postocular streak thin and dark (vs. light); G. scriptum ( Theobaldı 1868) by having one nasal (vs. two)ı 7 supralabials third and fourth in contact with the eye (vs. 8 supralabialsı 3rd and 5th in contact with the eye)ı lower number of subcaudals (67 – 73 vs. 87 – 98)ı dorsal scales in 15-15-15 rows (vs. 13-13-13 rows)ı postocular streak thin (vs. none).
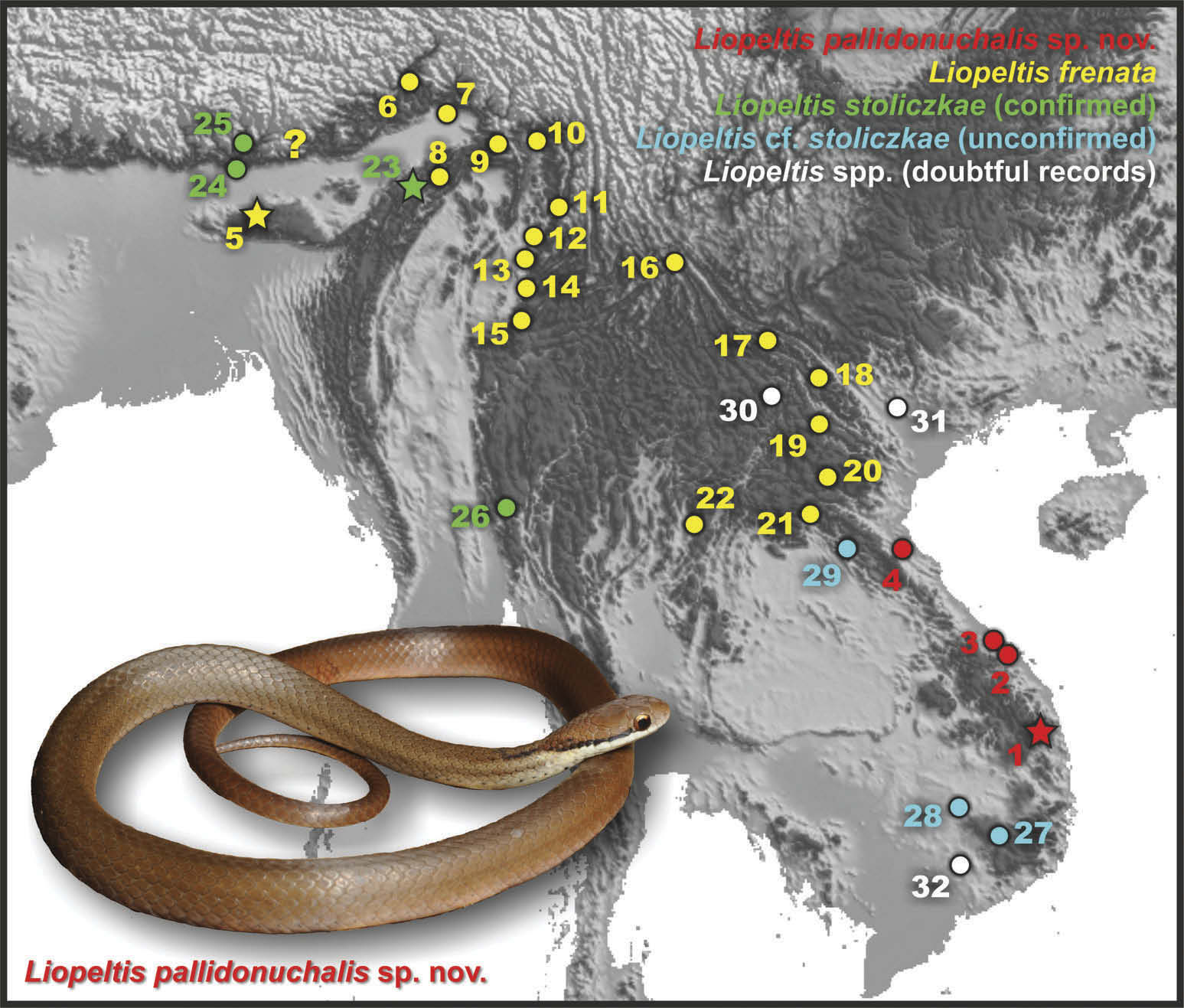







Table 2. Main morphological variation in the type series and referred materials (ref.mat.) of Liopeltis pallidonuchalis sp. nov. See Materials and Methods for other abbreviations.
| Number | ZMMU R – 15682 | DTU 307 | ZFMK 83105 | VNUH 9.7. ’ 06-1 |
|---|---|---|---|---|
| Type status | Holotype | Paratype | Paratype | ref.mat. |
| Sex | M | F | F | M |
| SVL | 297 | 275 | 336 | 270 |
| TaL | 112 | 111 | 130 | 116 |
| TL | 409 | 386 | 466 | 386 |
| Rel TL | 0.274 | 2.88 | 0.279 | 0.301 |
| DSR | 15-15-15 | 15-15-15 | 15-15-15 | 15-15-15 |
| VE | 2 + 133 | 2 + 138 | 2 + 138 | 4 + 126 |
| SC | 67 | 73 | 71 | 69 |
| Anal | 2 | 2 | 2 | 2 |
| SLb | 7 | 7 | 7 | 7 |
| SLE | 3 – 4 | 3 – 4 | 3 – 4 | 3 – 4 |
| ILb | 8 | 8 | 8 | 7 |
| Loreal | 0 | 1 | 1 | 1 |
| Praefrontal touching SLb | 1 | 0 | 0 | 0 |
| PrO | 1 | 1 | 1 | 1 |
| PoO | 2 | 2 | 2 | 2 |
| At | 1 | 1 | 1 | 1 |
| Pt | 2 | 2 | 2 | 2 |
| Source | This study | Ziegler et al. (2007) |
| ZMMU |
Zoological Museum, Moscow Lomonosov State University |
| R |
Departamento de Geologia, Universidad de Chile |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Liopeltis pallidonuchalis
| ı Tan Van Nguyen, Nikolay A. Poyarkov Jr. & Vogel, Gernot 2019 |
Liopeltis frenatus
| Ziegler Ti Hoang QX 2009: 115 |
| Ziegler Ti Hendrix Ri Vu NTi Vogt Mi Forster Bi Dang NK 2007: 10 |
| Smith MA 1943: 183 |