
Nigrobaetis rhithralis (Soldán & Thomas, 1983)
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4462.1.2 |
|
publication LSID |
lsid:zoobank.org:pub:3B6E40BD-BC67-471D-AC7B-B43DDC9917C4 |
|
DOI |
https://doi.org/10.5281/zenodo.5952356 |
|
persistent identifier |
https://treatment.plazi.org/id/03A087D1-FFB0-FFDC-FF6D-FA88122AFB95 |
|
treatment provided by |
Plazi |
|
scientific name |
Nigrobaetis rhithralis (Soldán & Thomas, 1983) |
| status |
|
Nigrobaetis rhithralis (Soldán & Thomas, 1983) View in CoL
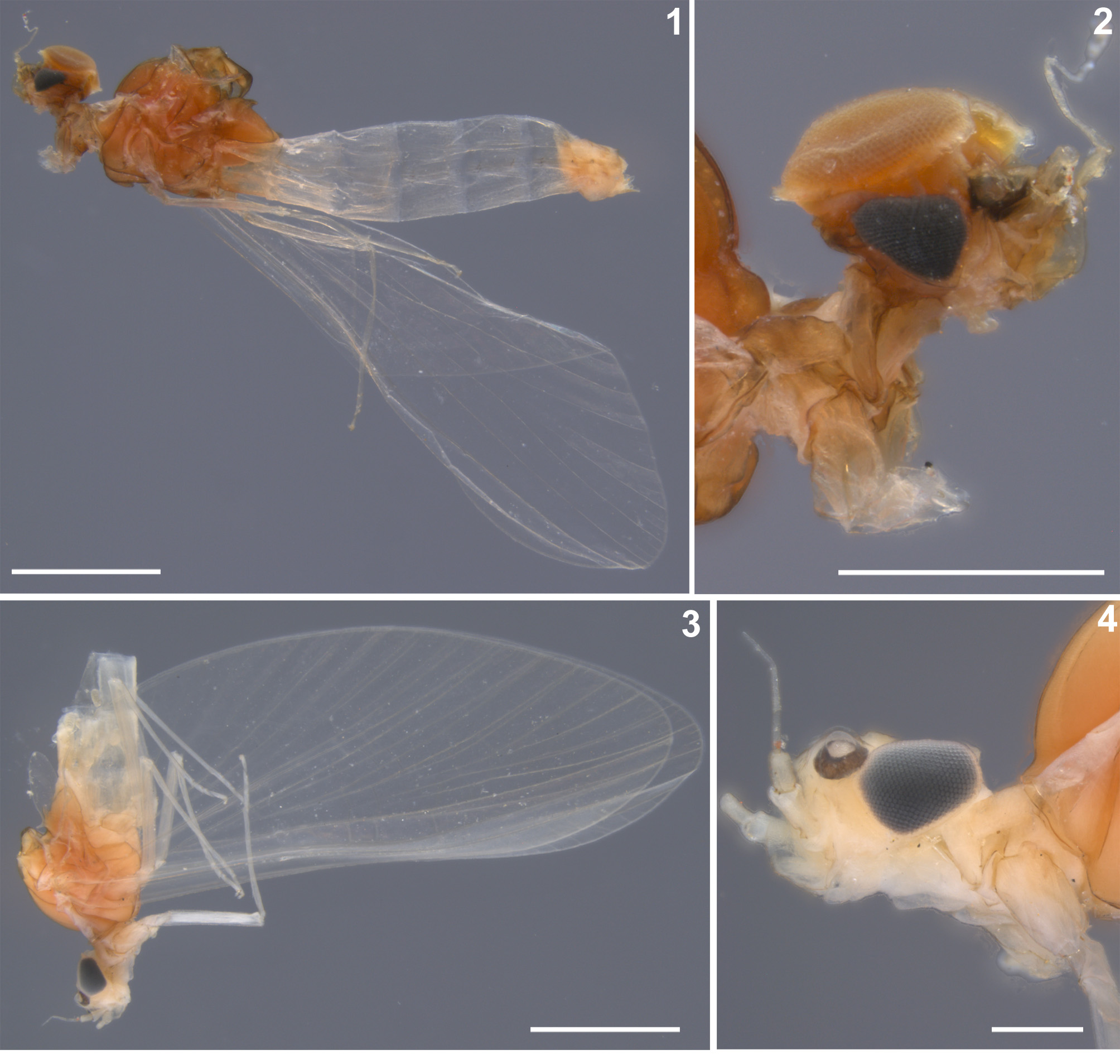
Figures 1–9 View FIGURES 1–4 View FIGURES 5–9 , 10A–B View FIGURE 10 , 11–60 View FIGURES 11–12 View FIGURES 13–16 View FIGURES 17–25 View FIGURES 26–35 View FIGURES 36–39 View FIGURES 40–48 View FIGURES 49–54 View FIGURES 55–60 , Table 1
Baetis rhithralis Soldán & Thomas, 1983 : Acta Entomol. Bohemoslov., 80: 356
? Baetis rhithralis Soldán & Thomas, 1983 : Novikova & Kluge, 1994: Entomol. Obozr., 53 (3): 632 [synonymy assumption; Nigrobaetis the subgenus of Baetis s.l.]
Diphetor rhithralis (Soldán & Thomas), 1989 [sic!]: Waltz et al., 1994: Bull. Soc. Hist. Nat., Toulouse, 130: 34 [new generic combination]
Diphetor rhithralis (Soldán & Thomas, 1983) : El Alami et al., 2000: Zool. Baetisca, 11: 110 [faunistics, new record from Morocco]
Nigrobaetis rhithralis (Soldán & Thomas, 1983) View in CoL : Thomas, 1998: Bull. Soc. Hist. Nat., Toulouse, 134: 17 [faunistics, formal new generic combination]; Zrelli et al., 2011: Rev. Suisse Zool., 118 (1): 7 [faunistics, new record from Tunisia]; Zrelli et al., 2012: 67; Zrelli et al., 2016: Inland Water Biology, Suppl. 3: 102
Nigrobaetis rhithralis (Soldán & Thomas) View in CoL : Lugo-Ortiz & de Moor, 2000: African Entomol., 8 (1): 72 [systematics; discussion on generic placement]
Baetis rhithralis: Jacob, 2003 : Lauterbornia , 47: 121 [species grouping; faunistics]
Baetis rhithralis Soldán & Thomas, 1983 : Schwarz-Waubke et al., 2003: Beitr. Naturk. Oberöster., 12: 409 [list of types]
Baetis (Nigrobaetis) rhithralis Soldán & Thomas, 1983 : Bauernfeind & Soldán 2012: The Mayflies of Europe ( Ephemeroptera ): 156, 574, fig. 69 [taxonomy, distribution, biology]
Type series. Original description of the species N. rhithralis is based on the male imago (holotype), female subimago and larvae (paratypes). The type series consists of 16 larvae, 3 imagos of both sexes (2 male imagos including the holotype, and one female imago), as well as 7 subimagos of both sexes (3 males and 4 females). All adults were reared from larvae, with preserved subimaginal and larval skins. Genitalia, forelegs and hind wings of the holotype and one male imago paratype were mounted on slides in Canada balsam and Liquid de Faure ; 3 larvae and 6 larval skins were mounted on slides in Euparal or Canada balsam. Other adults are partly damaged (see below).
Type locality can be defined as follows: Algeria, Wilaya de Blida [Blida Province], Chréa National Park, Blidean Atlas (part of the Tell Atlas), Camp des Chênes, Gorge de la Chiffa, confluence of Oued Chiffa [Chiffa River] and Ruisseau des singes, 250–270 m a.s.l., 36°24'04"N, 2°45'50"E, 09.x.1981, T. Soldán leg. (see Figs 82, 83 View FIGURES 82–83 ).
In the original description ( Soldán & Thomas 1983a), the authors erroneously indicated that the type material of N. rhithralis was collected in Oued Merdja. However, this stream (original name is Oued Merdja Kbira) is situated somewhat southern in upper part of Oued Chiffa. The same mistake was made with the type locality of B. (Rhodobaetis) chelif Soldán & Godunko, 2005 , as a part of the paratypes (adults and larvae) of this species was collected in the same place and date as the type series of N. rhithralis (see Soldán et al. 2005).
Investigated type material of N. rhithralis preserved in ethanol was originally labelled as follows: [ Baetis rhithralis Sold. & Thomas T. Soldán det. 1982]; [ALGER, Oued Merdja, Rouisseau des Singes 9.10.81 T. Soldán]. Place of deposition: IE (T. Soldán collection).
Other material: 1 larva, Tunisia, Bizerte, Joumine, Basin Ichkeul , Riv. Joumine ( Upstream dam); 57°46’09’’N 9°31’29’’E; 06.iv.2009; coll. Sonia Zrelli ( MZL collection). GBIFCH Code: GBIFCH00331449. GoogleMaps
Diagnosis. Nigrobaetis rhithralis can be distinguished from other representatives of the genus Nigrobaetis by the combination of following characters:
male imago: facetted surface of turbinate eyes intensively dark orange, shaft with two ring of different colour [according to original description]; faceted surface weakly ellipsoidal; shaft moderately high and clearly diverge after middle of its length; hind wings with two longitudinal veins ( Figs 7, 8 View FIGURES 5–9 ); second longitudinal vein forked at 0.48–0.56 of its length; abdominal terga I, VII–X rusty dark brownish; unistyliger nearly square-shaped, without apico-internal process; segment II of forceps with small inner hump; segment III of forceps elongated and rounded apically;
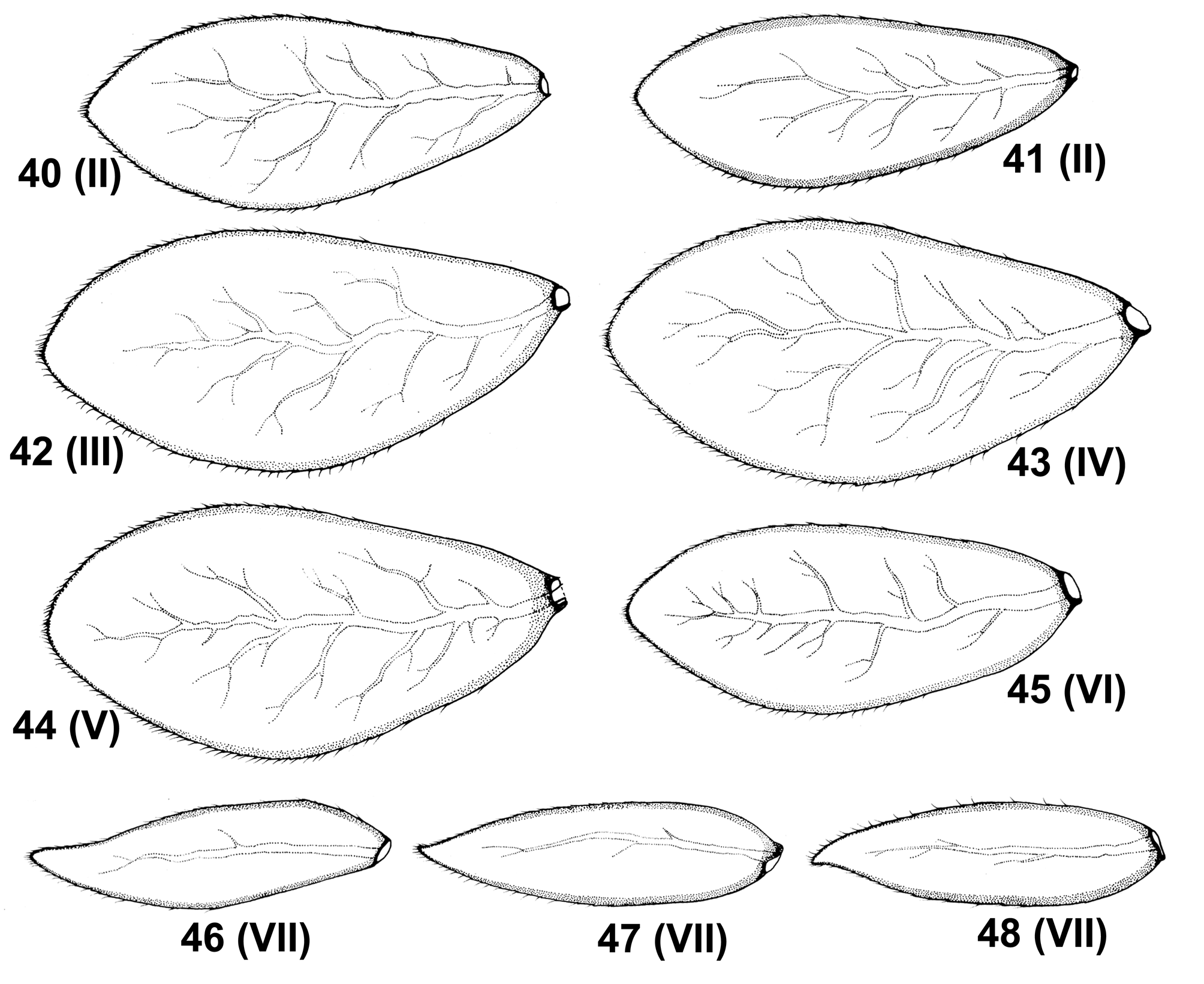
larva: surface of antennal segment I and II with scattered short setae ( Fig. 49 View FIGURES 49–54 ); flagellar segment with petalshaped scales and their sockets on surface, prominent slender spines on posterior margin; nearly U-shaped frontal suture ( Fig. 50 View FIGURES 49–54 ); labrum with 1+2 (sometimes up to 4) long submarginal setae ( Figs 17, 18 View FIGURES 17–25 ); subapical setae absent on glossae dorsally; segment III of labial palp oblong-shaped, with inner margin nearly straight or slightly concave; 9–12 stout setae with jagged margins along outer margin of femora, 22–26 on inner margin of tibia, and 14–25 on inner margin of tarsi; tarsal claw with 12–16 spines, without subapical tiny setae; posterior margin of abdominal terga ĪIII (rarely VI) with a row of relatively short, stout spines; surface of terga and legs (especially femora) with distinct dominance of petal-shaped scales; gill I absent, gills II–VI asymmetric, gill VII slightly asymmetric or nearly symmetric; posterior prolongation of paraproct absent; inner margin of paraproct and cercotractor with respectively 28–35 and 18–25 spines arranged in one row; transversal band absent on caudal filaments; terminal filament 1/2–2/3 of cerci; cuticular coloration of abdominal segments uniform, with a pair of paramedian pale strokes and spots.
Description. Male imago: Measurements: body length 4.2 mm; forewing length 4.5̄ 4.6 mm; cercus length 5.0– 5.2 mm. Specimens are partly damaged. The proportions of turbinate eyes are given in Table 1; for details see also Novikova & Kluge (1994: 630, table 3).
Due to long-term storage of the material in ethanol (more than 30 years), the colour pattern of male imagos and other adults of N. rhithralis slightly differs from those described by Soldán & Thomas (1983a) ( Figs 1–4 View FIGURES 1–4 , 11, 12 View FIGURES 11–12 ). Regarding the general colour pattern published in the original description, the body colour of historical material is faded. Cuticular coloration is poorly distinguishable, especially on abdominal segments.
Head. Vast part unicolorous light brown; darker smudges seen proximally. Antennae yellowish basally, yellowish-white apically. Dirty yellowish smudge seen between lateral ocelli. Ocelli yellow to dirty yellow apically, black basally. Turbinate eyes moderately high, well separated and diverge near to middle of their length ( Figs 2 View FIGURES 1–4 , 5, 6 View FIGURES 5–9 ); distance between basal part of turbinate eyes approximated equal to width of central ocellus; distance between upper parts of eyes at least three times more than base of central ocellus (for other proportions see Table 1). Faceted surface of turbinate eyes ellipsoidal dorsally ( Fig. 5 View FIGURES 5–9 ), slightly flattened; outer and inner margins distinctly convex. Faceted surface unicolorous, fadedly dirty brown [intensively dark orange: original description]. Colour pattern of turbinate eyes shaft poorly preserved ( Fig. 2 View FIGURES 1–4 ), i.e. apical strip only slightly paler than basal strip [broad paler strip apically and dark brown strip basally: orig. descr.].
Thorax. General colour pitch brown; pro- and metathorax slightly paler than mesothorax, light yellowishbrown to brown ( Fig. 1 View FIGURES 1–4 ). Ventral side of thorax paler than dorsal side, yellowish-brown.
Wings hyaline, translucent, unicolorous [milky grey: orig. descr.] ( Fig. 1 View FIGURES 1–4 ). Longitudinal venation of fore wings yellowish-white; cross venation pale, poorly visible; pterostigma slightly frosted-white, with 7–10 cross veins (several of them bifurcated). Hind wings elongated, 2.90–2.94 times longer than wide; costal process of hind wings well developed, not hooked ( Figs 7, 8 View FIGURES 5–9 ). Two well developed longitudinal veins present at hind wings, occasionally with short third vein near base of wing ( Fig. 8 View FIGURES 5–9 ). Second longitudinal vein forked at 0.48–0.56 of its length, with one intercalary vein in fork; second longitudinal vein fork asymmetrical.
Legs yellowish to dirty yellowish, with slightly darker femora [whitish, unicolorous: orig. descr.]. Mean length ratio of tarsomeres of foreleg as in Table 1 and Fig. 9 View FIGURES 5–9 .
Abdomen. Terga without visible pattern; segments II–VI uncoloured, translucent, with small greyish shadows anterolaterally [white, without markings: orig. descr.]; terga I, VII–X light brown, with inconspicuous light brown smudges [rusty dark brownish: orig. descr.].
Cerci unicoloured yellowish-grey, paler apically.
Genitalia. General colour yellowish to light brown; styliger, including unistyliger segments darker, forceps segments paler. Proportions of individual forceps segments slightly differ from those in original description: unistyliger nearly square-shaped ( Fig. 10A–B View FIGURE 10 ), only slightly tapered apically, without apico-internal process, approximately 0.78–0.82 times longer than wide; segment I conical, slightly narrowed towards apex [segment I oblong-shaped, about twice broader than long: orig. descr.]; segment II expanding apically, with small blunt hump on inner margin basally [segment II slightly tapered in middle, with inconspicuous bluntly pointed projection at the same place: orig. descr.]; segment III narrow and elongated, slightly expanding apically, rounded, 3.10–3.40 times longer than wide [2.5: orig. descr.].
Female imago. Measurements: body length 5.0 mm; forewing length 4.9̄5.0 mm; cercus broken. Specimen is partly damaged. General colour yellowish to yellowish brown, paler than in male imago ( Fig. 3 View FIGURES 1–4 ).
Head. Yellow, with darker smudges near eyes dorsally. Posterior margin of head nearly straight, only slightly convex medially ( Fig. 12 View FIGURES 11–12 ). Antennae yellow to greyish-yellow, scape darker. Eyes widely separated, unicoloured black, ocelli whitish apically, black basally ( Fig. 4 View FIGURES 1–4 ).
Thorax. Yellowish-brown to brown dorsally, yellow ventrally. Wings hyaline, translucent; forewings slightly frosted light yellow, with milky pterostigma ( Fig. 3 View FIGURES 1–4 ). Longitudinal veins of forewings yellow to whitish-yellow, paler in Cu–A field. Hind wings with venation similar to those in male imago. Legs unicoloured whitish-yellow, coxae slightly darker.
Abdomen. Terga whitish-yellow, partly translucent; segments VIII–X paler, yellow to dirty yellow.
Male subimago. Measurements: body length 4.3 mm; forewing length 4.4̄ 4.5 mm; cercus broken. Specimens partly damaged.
Head. Yellowish-brown to dirty brown; scape and pedicel brown; flagellum whitish-yellow; eyes and base of ocelli blackish; upper part of ocelli whitish. Faceted surface of turbinate eyes dirty brown; shaft pale, divided into two portions, with dirty yellow upper strip and light brown lower strip (margins between these both strips poorly visible).
Thorax. General colour light brown to brown; dark brown bands seen along of thoraxal sutures. Ventral side of thorax paler than dorsal side, yellowish-brown. Wings opaque, dirty yellowish; longitudinal venation yellow, darker basally; cross veins poorly distinguishable. Legs unicoloured whitish, forelegs same colour as other ones.
Abdomen. General colour brown, last two segments slightly paler; unclear pattern with one central and two oblique lateral strokes [a pair of divergent strokes: orig. descr.] seen medioanteriorly on terga II–VII (VIII). Sterna slightly paler than terga, yellow; longitudinal brownish smudges presented along of outer segment margins. Genitalia with yellow styliger and unistyliger segments, and whitish-yellow forceps segments.
Female subimago. Measurements: body length 4.8–5.0 mm; forewing length 5.0 mm; circus broken. Specimens are partly damaged.
General body colour ( Figs 11, 12 View FIGURES 11–12 ) similar to those of female imago; thorax slightly paler, yellowish-brown to light brown; abdominal segments unicolorous light brown, with pale pattern medially similar to those in female imago. Forewings colour similar to those of male subimago or slightly darker, frosted-yellow. Preserved part of cerci whitish, with yellow tip.
Mature nymph: Measurements: body length 4.2̄ 4.4 mm [male], 5.5̄ 6.2 mm [female]; cercus length 2.0̄ 2.4 mm.
Due to long-term storage of the material in ethanol, the colour pattern slightly differs from those described by Soldán & Thomas (1983a), i.e. more paler, faded. Cuticular colouration is poorly distinguishable in larvae ( Figs 13–16 View FIGURES 13–16 ), but relatively well preserved in larval skins mounted on slides.
Cuticular colouration. General colour yellowish-brown to brown. Head relatively pale, light yellow to yellowish-brown; distinct diffuse pale spot on clypeus and between base of antennae; diffuse pale maculation presented around of epicranial suture; genae yellow to light brown. Antennae generally unicolorous whitish-yellow to light brown, except slightly paler tip of flagellum. Surface of cuticula of larval turbinate eyes dirty yellow to light brown ( Fig. 15 View FIGURES 13–16 ). Labrum yellow to light brown, darker basally; mandible yellow to light brown.
Thorax yellow to brown; pronotum with a pair of diffuse oblique stokes; several diffuse whitish to yellow spots on meso- and metanotum dorsally; distinct yellowish spots near forewing pads base; forewing pads yellowish to light brown, with pale smidges over whole surface; hind wing pads unicolorous, light brown, with paler smudges basally. Thoraxal pleurae and sterna paler than terga, whitish-yellow to light brown, slightly darker around sutures.
Legs whitish-yellow to yellow ( Fig. 13 View FIGURES 13–16 ). Inconspicuous diffuse yellowish spot seen centrally, and yellow to light brown band located along inner and outer margins of each of femora; tibiae and tarsi unicolorous yellow proximally and centrally, slightly darker distally; tarsal claws light brown.
Abdominal segments yellowish-brown to brown. Terga with poorly distinguishable pattern. Tergum I with diffuse yellowish spots centrally and laterally. Terga II–VII with a pair of pale oblique divergent strokes ending by a small pale spots centrally; diffuse pale spots seen along anterior and lateral sides of segments. Terga VIII–IX pale, with broad diffuse spot centrally; inconspicuous smudges situated anteriolaterally. Tergum X with pale spot surrounded by a brownish smudge centrally. Abdominal sterna unicolorous yellow to yellowish-white, without distinct pattern. Terminal filaments unicolorous yellow to yellowish-brown, without transversal darker band centrally.
Hypodermal colouration. Head and thorax with light brown maculation dorsally and laterally. Faceted surface of larval turbinate eyes dirty yellow, light brown or dirty orange; shaft of larval turbinate eyes with more or less distinguishable two portions, i.e. upper dirty yellow to whitish-yellow and lower yellow to light brown. Legs yellow; small brownish spot seen near distal end of each femora and tibiae. Abdominal terga yellow to yellowishbrown; sterna unicolorous yellow. Ventral ganglionic chain not distinguishable. Gills whitish-yellow to dirty yellow; tracheae dirty violet; jagged margins of gills poorly visible, yellowish-brown coloured.
Head. Frontal suture nearly U-shaped ( Fig. 50 View FIGURES 49–54 ); surface of frons and clypeus covered with occasional B and Hr; more abundant on genae. Antennae up to 3 times longer than head ( Fig. 13 View FIGURES 13–16 ). Surface of flagellar segments with a few small SC-pet and SCS-pet (concentrated mainly near distal edge of segment), sparse Hr and B ( Fig. 29 View FIGURES 26–35 ). Posterior margin of flagellar segments with dense row of 21–30 well separated, relatively slender, bluntly pointed apically spines of different size (cuticular lobes sensu Gaino & Rebora 1999a: 70, figs 13a, b; 1999b), alternating with 1 or grouped by 2 B setae. Scape and pedicel with occasional small SC-pet and SCS-pet, and densely scattered Hr and B setae.
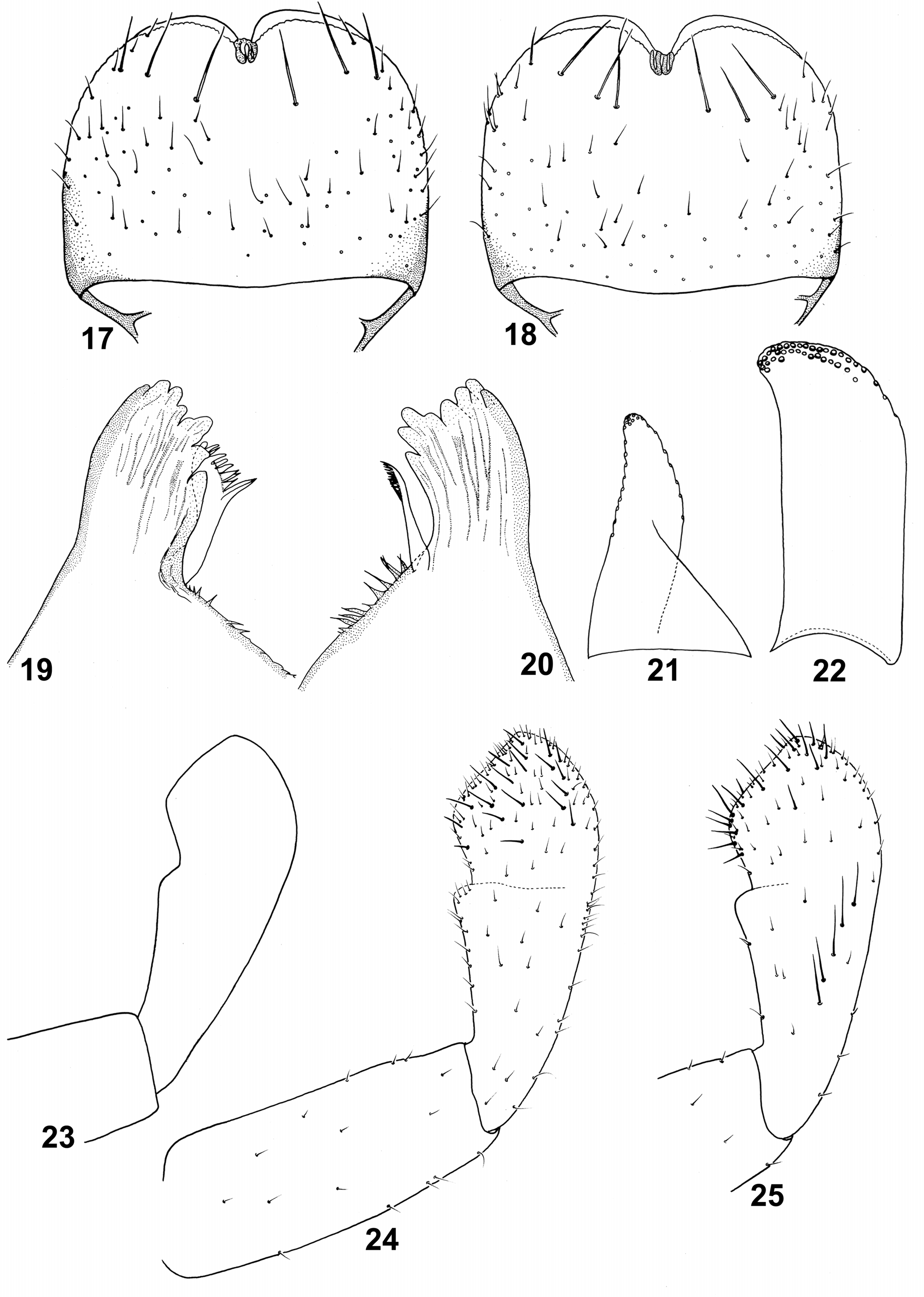
Mouthparts. Labrum ( Figs 17, 18 View FIGURES 17–25 ) relatively slender, tarring distally, with width/length ratio 0.41–0.45; dorsal surface with 1+2–4 long submarginal setae, concentrated more laterally [1+3–4 bristles: orig. descr.]; outer margin with one short seta; elongated Hr on labrum surface.
Hypopharynx and maxillae typical for Nigrobaetis , without any distinguishing features.
Left mandible ( Fig. 19 View FIGURES 17–25 ) with 7 incisors rounded apically; prostheca with 11–13 small spines (2–3 basal spines bluntly pointed, longer than other ones; 8–10 rounded and short). Right mandible ( Fig. 20 View FIGURES 17–25 ) with 8 incisors rounded apically; prostheca slender, not distinctly extended apically, with numerous pointed setae. Both mandibles with slim pointed scales between prostheca and mola (see Table 1); 17–24 fine setae between prostheca and basal incisor of right mandible.
Labium with glossae shorter than paraglossae. Glossae conical ( Fig. 21 View FIGURES 17–25 ), relatively wide, slightly curved, with
10–11 stout bristles along inner margin; 9–10 stout bristles on outer margin; surface covered with sparse B, short and long Hr setae. Paraglossae ( Fig. 22 View FIGURES 17–25 ) with two irregular rows of stout bristles apically on dorsal surface; 4–5 medium bristles in one row ventrally.
Shape of labial palp segments II–III as in Figs 23–25 View FIGURES 17–25 ; segment II with small, nearly rounded apicomedial projection, slightly expanded laterally; dorsal surface of segment II with row of 5–6 long setae centrally; segment III asymmetrical, covered with sparse stout setae and Hr dorsally and ventrally.
Thorax. Surface of thorax covered with short Hr, occasional B and SCS-pet.
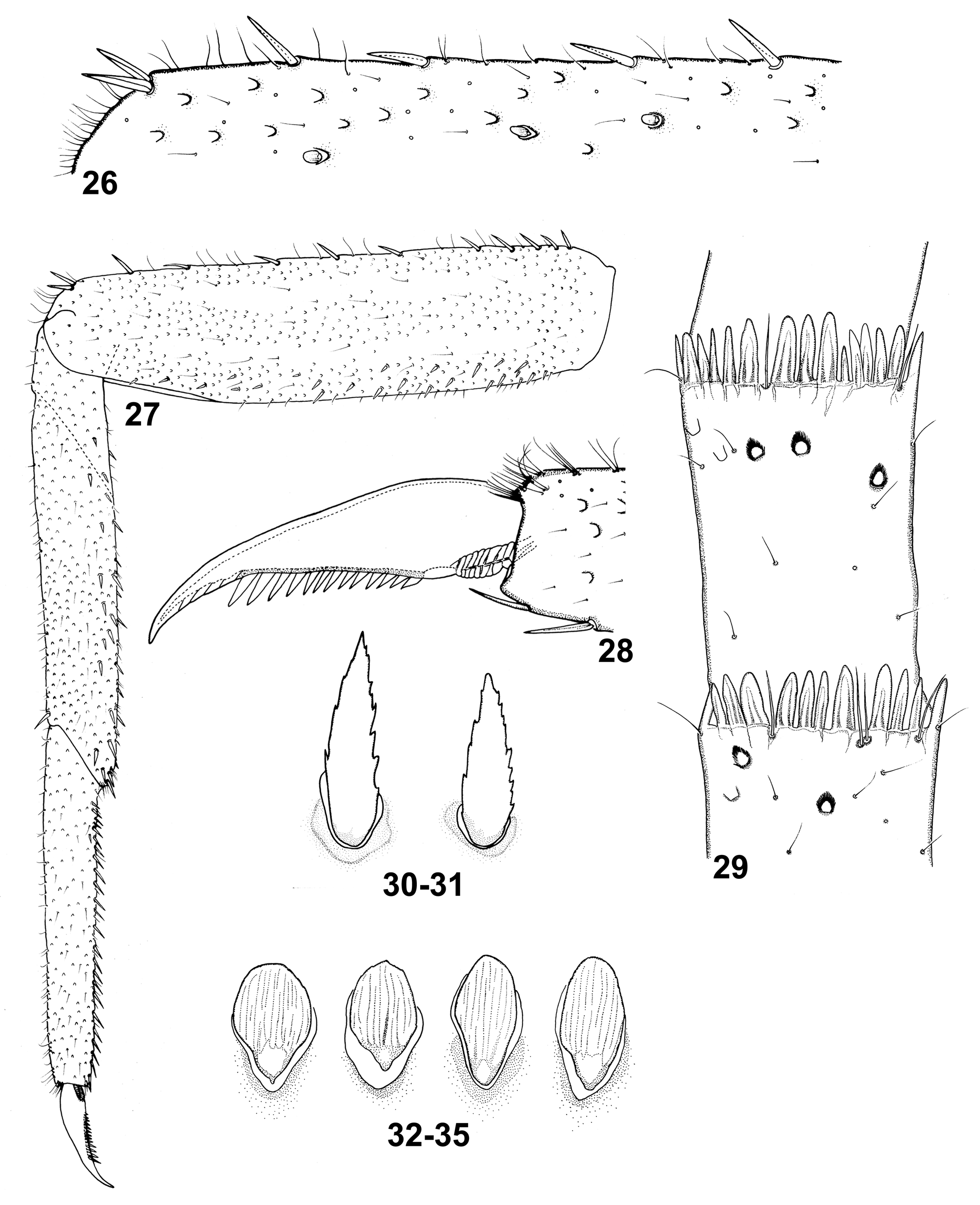
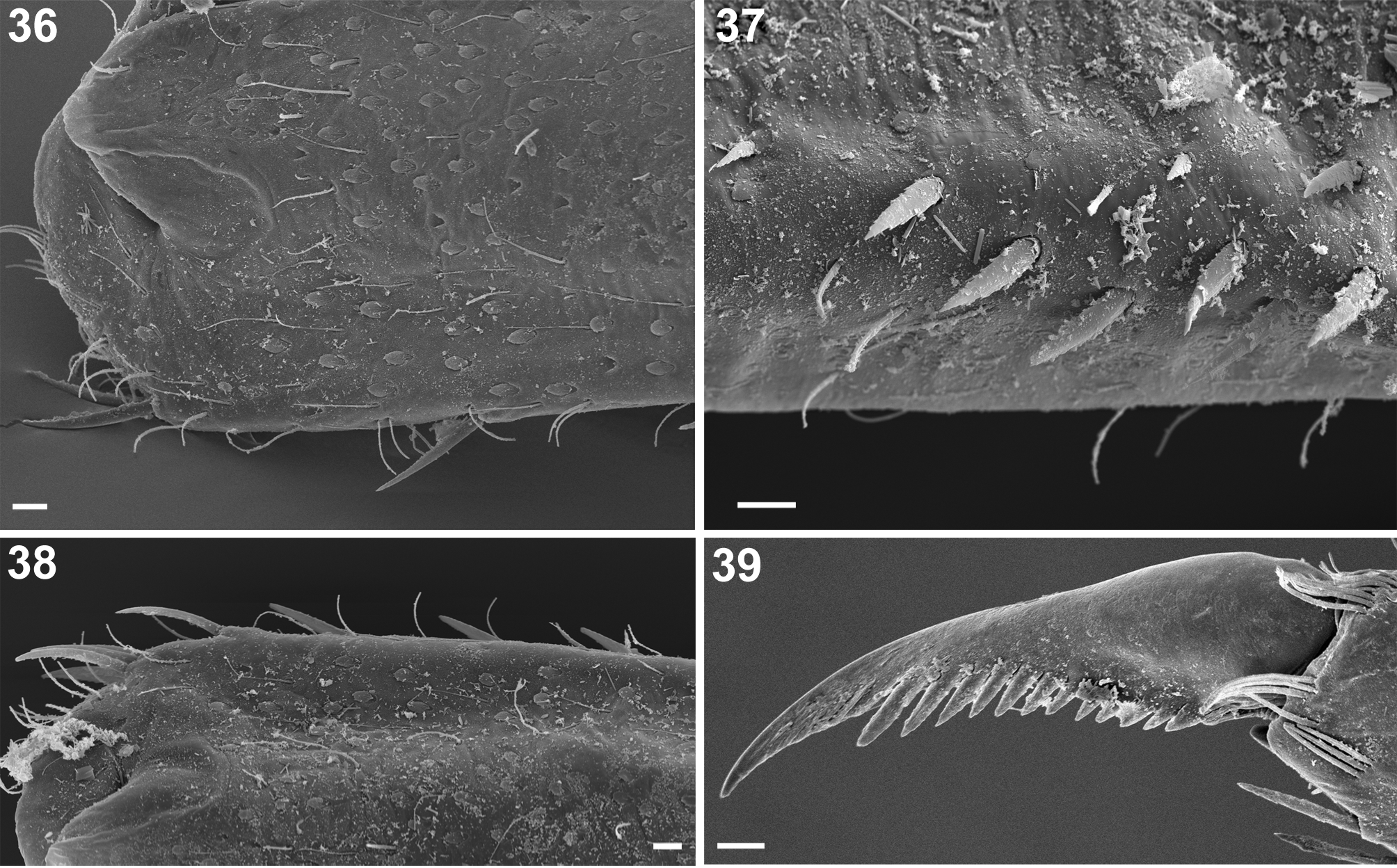
Forelegs. Irregular sparse row of 9–12 finely jagged DS-STS-bp and occasional DS-STS-p seen along outer margin of each of femora dorsally ( Figs 26, 27 View FIGURES 26–35 , 38 View FIGURES 36–39 ), alternating with Hr; outer margin with group of 2 (rarer 3–4) such setae on distal edge; submarginal field with sparse Hr, most abundant SCS-pet, and only a few small SC-pet and B setae. Dorsal surface of femora with similar types of setae ( Figs. 26 View FIGURES 26–35 , 36 View FIGURES 36–39 ); numerous SC-pet (dominant) and their bases can be concentrated along lateral margins and centrally (as a relatively distinct strip), or nearly evenly and densely scattered over whole surface. Inner margin and submarginal field of femora with 20–28 clearly jagged margins, small and middle DS-STS-p and -bp ( Fig. 37 View FIGURES 36–39 ). Outer margin of tibia generally with 0–7(8) finely jagged margins, middle and elongated DS-STS-p ( Fig. 27 View FIGURES 26–35 ); inner margin and submarginal field with 22–26 clearly jagged on margins setae of same type and Hr, and with groups of 2–3 B setae. Surface of tibia with Hr of different length, a few small DS-STS-bp and B, and dominant SCS-pet. Outer margin of tarsi covered with sparse Hr; inner surface with 14–25 clearly jagged middle DS-STS-p and - bp, alternating with 1 or grouped by 2–3 B setae. Surface of tarsi with occasional small SC-pet, and more abundant Hr and SCS-pet ( Fig. 27 View FIGURES 26–35 ). Tarsal claw elongated, tip slightly hooked and narrow; 12–16 teeth increasing in length towards claw apex ( Figs 28 View FIGURES 26–35 , 39 View FIGURES 36–39 ).
Abdomen. Posterior margin of tergum I with short stout spines of different size concentrated centrally, alternating with B and long Hr; terga II–III(VI) with a row of shortening spines approximately of same size, alternating with B and long Hr ( Figs 55, 56 View FIGURES 55–60 ); terga V–X with elongated stout spines, B and long Hr ( Figs 57–59 View FIGURES 55–60 ). Marginal spines with wide basis, sharply pointed apically, slightly asymmetrical, their lateral sides irregularly concave. Surface of terga with numerous SC-pet (dominant), less abundant SCS-pet, long Hr, B, STSs-ov and STSssp ( Figs 51, 52 View FIGURES 49–54 , 55–59 View FIGURES 55–60 ); occasionally, high concentration of SCS-pet on segments surface. Ventral ganglionic chain not visible. Posterior margin of sterna with elongated, slender spines, pointed or bluntly pointed apically, and a few Hr and B setae. Surface of sterna with dominant SCS-pet, sparse SC-pet (concentrated near segment margins), Hr and B setae.
Gill I absent. Gills II–VI asymmetric with blunt or bluntly pointed tip ( Figs 40–48 View FIGURES 40–48 ); length/width ratio of gills II, IV and VI as in Table 1. Gill VII slenderer with slightly concave inner margin near apex, or leaf-shaped (nearly symmetric), pointed or bluntly pointed at the tip, 3.36–3.67 times longer than broad ( Figs 46–48 View FIGURES 40–48 ). Surface of gills covered with small SC-pet, occasional SCS-pet and Hr along both margins of gill; other surface with solitary long Hr and B. Both gill margins with B setae, inserted in small not articulated setal bases. Tracheation well distinguishable, yellowish to light dirty brown.
Inner margin of paraproct with 28–35 stout spines gradually increasing in size and arranged in one row ( Fig. 60 View FIGURES 55–60 ); absence of posterior prolongation of paraproct; surface of paraproct with numerous SCS-pet, and only a few SC-pet, B and long Hr mainly centrally. Cercotractor with SCS-pet and SC-pet, margin with 18–25 spines arranged in one row.
Terminal filaments structure as on Fig. 60 View FIGURES 55–60 and Table 1.
Discussion. As already mentioned, N. rhithralis and N. digitatus are rather similar. Differential characters of adults and larvae are listed in Table 1; main differences between these two species are marked with gray. Detailed study of material of N. digitatus from different areas ( Germany, Balkans, Pontic region, Cyprus and Caucasus) revealed minute differences with specimens from Sweden. Thus, morphological characters corroborate the preliminary molecular results (Project FREDIE: http://www.fredie.eu/, S. Rutschmann personal communication) and seem to indicate that the present concept of N. digitatus corresponds in fact to a complex of sibylline species. Consequently, for the comparison with N. rhithralis we used only the Scandinavian specimens ( Sweden and Finland). When specimens from other territories are also considered, we clearly indicate them as N. digitatus s.l.
The colour pattern of male and female adults of N. digitatus and N. rhithralis is only partly similar. These two species clearly differ in the colour of male turbinate eyes of freshly fixed material, i.e. (i) yellowish-brown to uniform reddish or blood-red at facetted surface and eyes shaft in N. digitatus (fresh material observation); (ii) intensively dark orange at faceted surface with two varicoloured rings on shaft in N. rhithralis ( Table 1).
Other differences in turbinate eyes of male imagos are as following: (i) in lateral aspect the eyes shaft of N. digitatus distinctly divergent directly from the base ( Figs 61–64 View FIGURES 61–64 ); (ii) in contrast to moderately high eyes shaft of N. rhithralis , well separated, clearly diverges after middle of its length ( Figs 2 View FIGURES 1–4 , 6 View FIGURES 5–9 ; Table 1: character II.2).
Visible differences are observed in the shape of faceted surface, i.e. (i) ellipsoidal and distinctly asymmetrical faceted surface in N. digitatus ; (ii) in contrast to more rounded and not stretched laterally in N. rhithralis ( Figs 62, 64 View FIGURES 61–64 and character II. 1 in Table 1). Described above shape and proportions of turbinate eyes correspond to respective indexes proposed by Novikova & Kluge (1994) (see Table 1: characters III.1–4).
Additionally, abdominal terga of adults of N. rhithralis are slightly darker than in N. digitatus ( Table 1: character VI.1). There are no clear differences between both species in the structure of fore- and hind wings venation. The hind wings venations of N. digitatus present some variability, especially regarding the place of bifurcation of the second vein relatively to their length, and the number of intercalaries in this fork ( Novikova & Kluge 1994: 632–634, fig. 3.13–17). The variability within N. rhithralis remains unknown ( Figs 7, 8 View FIGURES 5–9 and tabular characters V.1–5).
The male genitalia of N. rhithralis differs from that of N. digitatus by the shape and proportions of forceps segments (see our Fig. 10A–C View FIGURE 10 , Table 1: characters VII.1–7; Bauernfeind & Soldán 2012: 148, 157, 573 574, figs 0 62, 069): (i) a distinct difference between these species is found in the shape of the unistyliger with predominant presence (in N. digitatus ) or absence (in N. rhithralis ) of apico-internal process; (ii) in the shape and proportion of unistyliger and segment III; and (iii) in the shape of inner margin of segment II, with presence (in N. rhithralis ) or absence (in N. digitatus ) of a small but distinct inner hump.
The male imago reared from larva by Nikita J. Kluge (Psekups River, Russian Federation) and drawn by us ( Fig. 10C View FIGURE 10 ), was initially attributed to the species B. digigracilis [nomen nudum] based on features listed in Novikova (1987) (see also Novikova & Kluge 1994: 634). The shape and proportions of forceps segments of the specimen from Psekups River are rather similar to those in N. rhithralis ( Figs 10A–B View FIGURE 10 ). However, within males collected by us in closely located Tuapse River, forceps structure is the same as in other European representatives of N. digitatus s.l. ( Figs 65–67 View FIGURES 65–67 ). It should be emphasized, that differences of Caucasian material of N. digitatus s.l. as well as variability of characters of this species (especially in larvae), were described and discussed in details by Novikova & Kluge (1994).
The females of both species can be distinguished by the shape of posterior margin of head, markedly incised centrally in N. digitatus , and nearly straight in N. rhithralis ( Figs 11, 12 View FIGURES 11–12 ; Table 1: character VIII.1).
Larval differences between N. rhithralis and N. digitatus can be found in the colour pattern of body, especially of abdominal terga ( Table 1: characters IX.1, XI.1 and XII.1). Larval colour pattern in the specimens of N. digitatus s.l. from different territories (Ural, European part of the Russian Federation and Caucasus, Cyprus, Pontic Region) is rather variable (see also Novikova & Kluge 1994). Bauernfeind & Soldán (2012) summarized information on the main colour pattern of abdominal terga I–XI of N. digitatus s.l. and proposed two types, i.e. (i) a pair of large pale oblique spots or small dots and dashes centrally on light brown to brown background; (ii) the same pattern, in combination with predominantly pale terga IX–X [according to Novikova & Kluge (1994: 632, fig. 3.1) terga I, IV, VIII–X can be lighter, without pale spots on dark background] (for details see also Table 1). In contrast, N. rhithralis could be characterized by the uniform colour pattern of abdominal terga in all reinvestigated larvae and larval skins. This pattern markedly differs from that described and depicted for N. digitatus s.l. by Müller-Liebenau (1969: 171–172, figs 133, 134), Novikova & Kluge (1994: 632, fig. 3.1), Bauernfeind & Soldán (2012: 148), and observed for the North European specimens ( Table 1: VII.1 and Remarks for Table 1). Additionally, there is no transversal band on caudal filaments in N. rhithralis , in contrast to more or less visible, relatively narrow band in N. digitatus s.l. ( Table 1: character XII.20). Both species are also well distinguishable by the colouration of head and pronotum ( Table 1: characters IX.1 and XI.1).
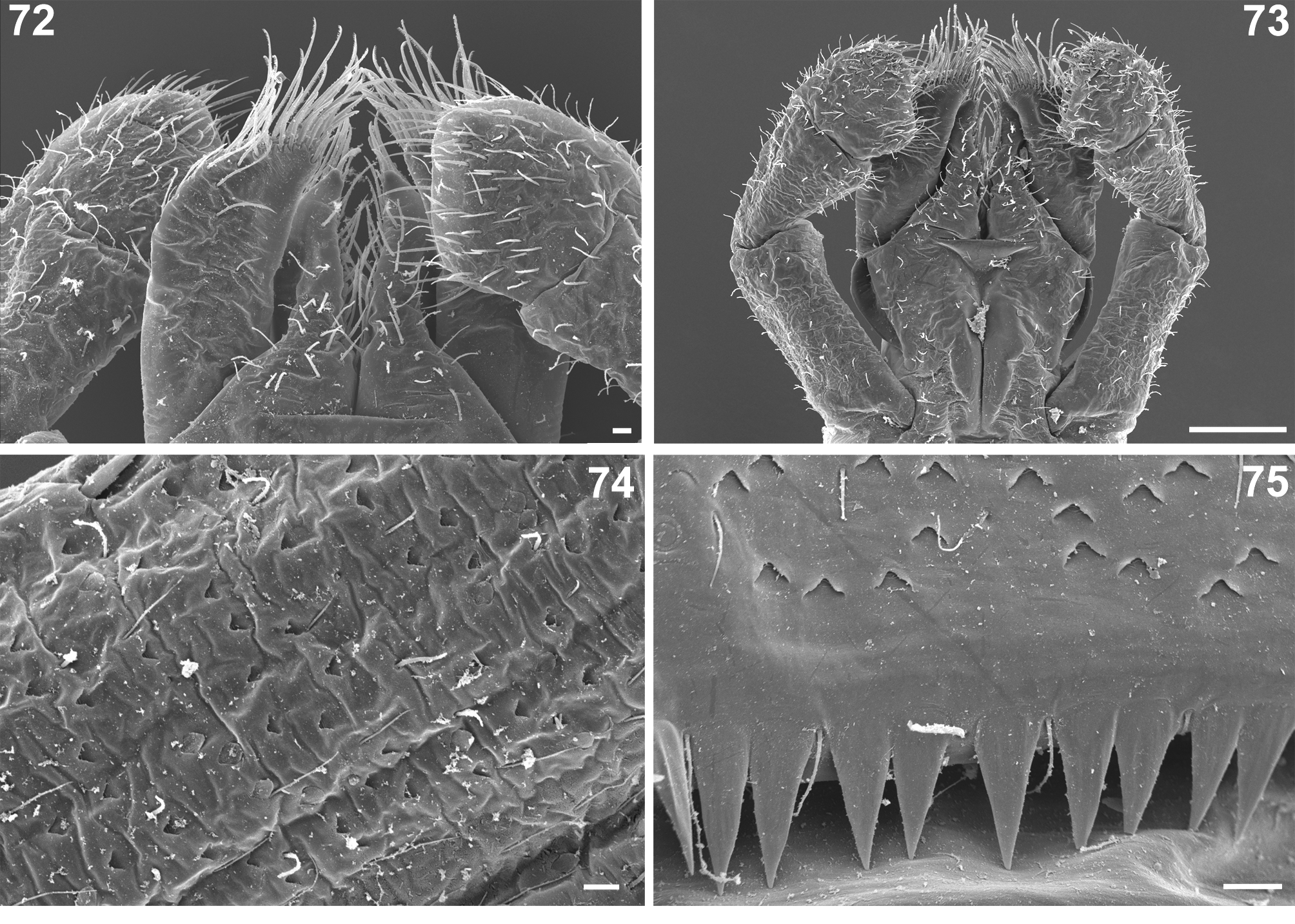
Nigrobaetis rhithralis and N. digitatus present similar setation of the body surface. However, the surface of antennae is more densely covered by setae and scales in N. rhithralis ( Figs 49 View FIGURES 49–54 , 68 View FIGURES 68–71 ; Table 1: character IX.2). The posterior margin of flagellar segment in this species is densely covered by relatively slender spines, in contrast to more robust spines in N. digitatus with different grouping of setae ( Figs 29 View FIGURES 26–35 , 69 View FIGURES 68–71 ; tabular character IX.4). Additionally, we observed a significant concentration of SC-pet setae on the surface of legs (especially femora) and on abdominal terga in N. rhithralis , in contrast to dominance of scale sockets in most specimens of N. digitatus ( Table 1: character XII.2). Both species also differ in the shape of posterior margin of abdominal terga ( Table 1: characters XII.3–5), i.e. by the shape and arrangement of marginal spines ( Figs 52 View FIGURES 49–54 , 55–59 View FIGURES 55–60 for N. rhithralis ; Figs 75 View FIGURES 72–75 , 78, 79 View FIGURES 76–81 for N. digitatus ).
Setation of the legs differs only by the number of pointed stout setae with jagged sides along outer margin of femora, and can be used for the delimitation of both discussed species ( Table 1: character XI.6). Quantitative values of other characters ( Table 1: XI.7–11) significantly vary in both species and are mostly overlapping. There are no clear differences in mouthpart structures between the two species, larvae of N. rhithralis only slightly differ from larvae of N. digitatus by some features in mandibles, glossa and paraglossa ( Table 1: characters X.5, 6, 14, 15).
……continued on the next page
……continued on the next page
……continued on the next page
……continued on the next page
Remarks
(1) A — coding according to Soldán & Thomas (1983a: 356–358) and based on type material observation; B — Eva Engblom personal communication (due to long-term storage in ethanol the turbinate eyes color of N. digitatus is faded and differs markedly from the original color observed in fresh material; see Figs 61–64 View FIGURES 61–64 ); C — based on fresh material observation.
(2) Indexes of male turbinate eyes proportions for N. rhithralis are coding based on type material reinvestigation.
A — maximal distance between outer margins of facetted surfaces; B — minimal distance between inner margins of facetted surfaces of turbinate eyes; C — length of facetted surfaces of turbinate eyes; D — height of turbinate eyes [distance between upper margin of eyes and upper part of faceted surface]; N — maximal distance between outer margins eyes; α — angle between the longest axis of facetted surfaces of turbinate eyes (see Novikova & Kluge 1994).
(3) See details and discussion in Müller-Liebenau (1965: 84; 1969: 171) for N. digitatus ; additional information in Novikova & Kluge (1994: 632–634, fig. 3:13–17), and Bauernfeind & Soldán (2012: 148).
(4) See Novikova & Kluge (1994: 630, key for male imagos) for N. digitatus ; Bauernfeind & Soldán (2012: 157) for N. rhithralis .
(5) Not shown for N. digitatus e.g. on fig. 1a in Müller-Liebenau (1965: 84) and Jacob (2003: 112, fig. 26b), but well visible on photo published by Müller-Liebenau (1969: 164, fig. 124).
(6) In N. digitatus separation of forceps segments I and II is indistinct (see e.g. Bauernfeind & Soldán 2012: 148); marked constriction at the junction of segments I and II according to Elliot & Humpesch (1983: 34, fig. 17c, in key for male imagines is marked as segment II and III).
(7) See Soldán & Thomas (1983a: 356–358, fig. 12) and Bauernfeind & Soldán (2012: 148, 157, figs 0 62, 069).
(8) Borrowed from Novikova & Kluge (1994: 633, fig. 3:11–12).
(9) A — based on original description ( Soldán & Thomas 1983a: 356–358) and type material observation; B — based on material observation.
(10) Depicted and discussed for several representatives of B. muticus species-group sensu Müller-Liebenau (1969) by Sartori & Thomas (1991: 226, 231, fig. pl. 3).
(11) Described and figured by Müller-Liebenau (1965: 87, fig 2b, c) for N. rhithralis ; description of setation and scanning electron photographs of setae published by Gaino & Rebora 1999b: 70–74, fig. 13a) for two species of the genus Nigrobaetis .
(12) For Nigrobaetis and other taxa of Baetinae the character is used by Novikova (1987) and Novikova & Kluge (1987, 1994); another character, viz. the shape of frontal keel is constant in all species of Nigrobaetis (incl. Takobia ) according to Kluge & Novikova (2014: 230).
(13) Character coded as in Bauernfeind & Soldán (2012).
(14) The character depicted by Müller-Liebenau (1965: 87, 2j) for N. digitatus , and by Soldán & Thomas (1983a: 358, fig. 2) for N. rhithralis ; stated for Nigrobaetis (incl. Takobia ) by Novikova & Kluge (1987: see tabular characters on the page 9); individual variability of this character tested by Kazancı & Thomas 1989: 323–324, fig. 1a–d); for five species belongs to the B. muticus species-group analysed by Zrelli et al. (2012: 66, table 1; sub. the genus Alainites ); for Indobaetis the character tested by Kluge & Novikova (2014: 230).
(15) Discussed in details by Kluge & Novikova (2014: 230).
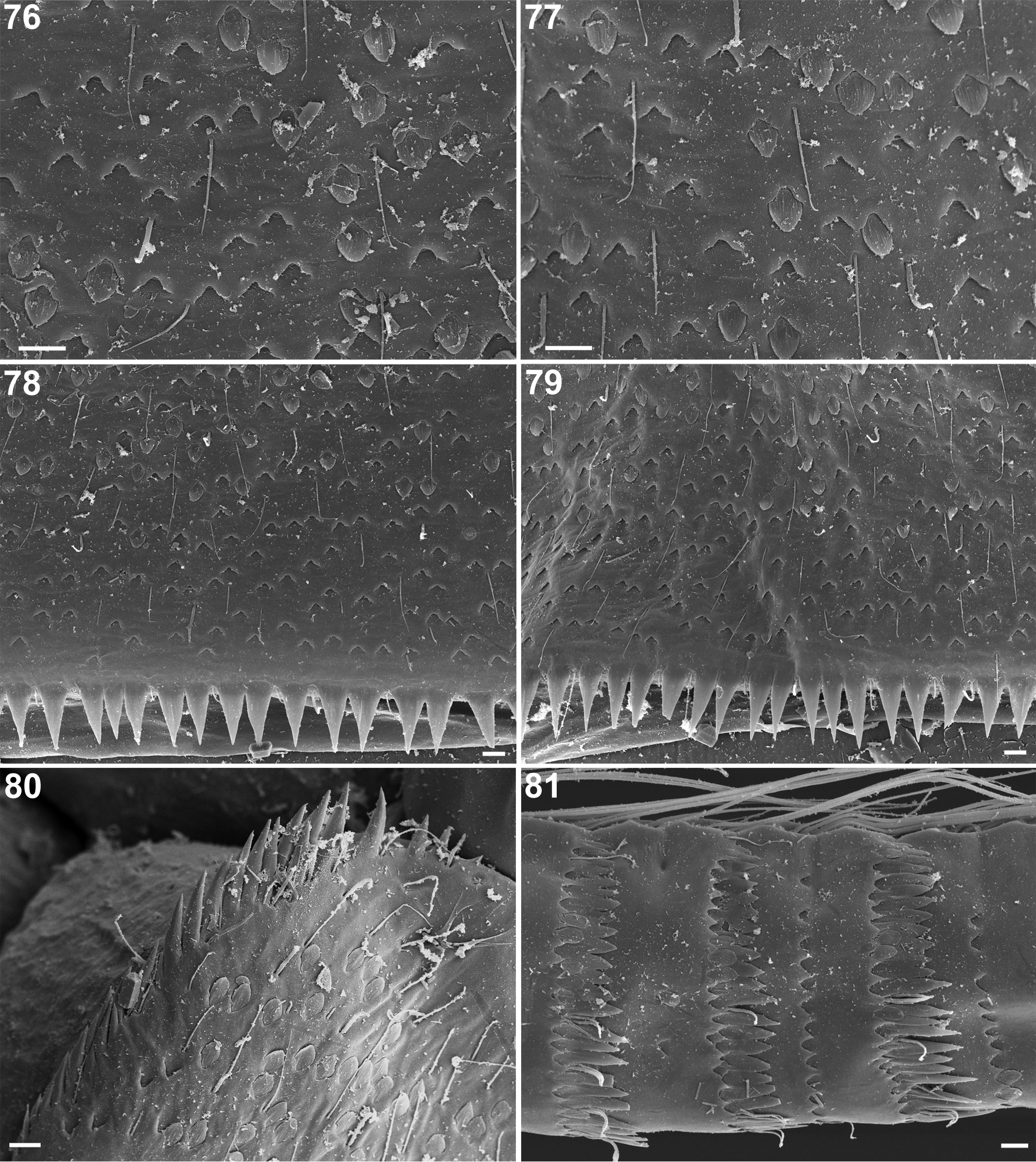
Differences can be also found in gill shape and their proportions ( Table 1: characters XII.10–13). Gills VII of N. digitatus possess a rather unique concave margin ( Müller-Liebenau 1969: 169, fig 132) while gills VII are almost symmetrical in N. rhithralis . The structure of lateral margins and surface of gills are rather similar in both species.
The larvae of N. rhithralis also markedly differ from larvae of N. digitatus by the combination of the following characters: (i) shape of frontal suture; (ii) spines number and arrangement along the inner margin of paraproct and cercotractor (XII.15 and XII.18; Figs 60 View FIGURES 55–60 , 80 View FIGURES 76–81 ); and (iii) shape of posterior margin of caudal filaments (XII.21; Figs 54 View FIGURES 49–54 , 81 View FIGURES 76–81 ).
No differences in eggs chorionic surface were found between the two species (for N. digitatus s.l. see Kopelke & Müller-Liebenau 1981: 3, figs 18, 19).
As N. digitatus View in CoL , N. rhithralis View in CoL possesses six pairs of gills, and thus differs from N. vuatazi Gattolliat & Sartori, 2012 View in CoL (described from Jordan) and the Afrotropical species N. harasab (Soldán, 1977) View in CoL , both with seven pairs of gills. From other Nigrobaetis View in CoL species described from the Afrotropical Region (viz., N. colonus Gattolliat, 2004 View in CoL , N. cryptus Gattolliat, 2004 View in CoL and N. bethunae Lugo-Ortiz & de Moor, 2000 ), N. rhithralis View in CoL markedly differs by the presence of hind wing pads ( Gattolliat 2004, Gattolliat et al. 2012). Nigrobaetis rhithralis View in CoL can be distinguished from the Algerian species N. numidicus (Soldán & Thomas, 1983) View in CoL and N. arabiensis Gattolliat & Sartori, 2008 View in CoL described from United Arab Emirates by the well-developed numerous spines along the inner margin of paraproct ( Soldán & Thomas 1983b; Gattolliat & Sartori 2008; Gattolliat et al. 2012). The second longitudinal vein of hindwings in N. numidicus View in CoL and N. vuatazi View in CoL is not bifurcated as in most species of the genus. Nigrobaetis harasab View in CoL , N. numidicus View in CoL and N. vuatazi View in CoL can be separated from N. rhithralis View in CoL and N. arabiensis View in CoL by the shape of spatulated stout setae along the outer margin of femora.
Distribution. The distribution of N. rhithralis is restricted to the Blidean Atlas as a part of the Tell Atlas in Algeria (see above; Soldán & Thomas 1983a) ( Figs 82, 83 View FIGURES 82–83 ); it is also known from two localities within the Rif Mountains in Morocco ( El Alami et al. 2000); and, according to the latest data, this species was also found in five localities in Northern Tunisia ( Zrelli et al. 2011, 2016).
Biology. See Soldán & Thomas (1983a) and Bauernfeind & Soldán (2012).










| MZL |
Musee Zoologique |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Nigrobaetis rhithralis (Soldán & Thomas, 1983)
| Godunko, Roman J., Martynov, Alexander V. & Gattolliat, Jean-Luc 2018 |
N. digitatus
| Godunko & Martynov & Gattolliat 2018 |
Nigrobaetis
| Godunko & Martynov & Gattolliat 2018 |
N. vuatazi
| Gattolliat & Sartori 2012 |
N. vuatazi
| Gattolliat & Sartori 2012 |
N. vuatazi
| Gattolliat & Sartori 2012 |
N. arabiensis
| Gattolliat & Sartori 2008 |
N. arabiensis
| Gattolliat & Sartori 2008 |
N. colonus
| Gattolliat 2004 |
N. cryptus
| Gattolliat 2004 |
Baetis rhithralis:
| Jacob 2003 |
N. bethunae
| Lugo-Ortiz & de Moor 2000 |
Baetis rhithralis Soldán & Thomas, 1983
| Soldan & Thomas 1983 |