
Blattochaeta peterhlavaci D. Čeplík, Lakota & J. Čeplík, 2021
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4969.1.4 |
|
publication LSID |
lsid:zoobank.org:pub:B0AFAF8C-B2A9-41FB-AE9C-07DE268EF82E |
|
DOI |
https://doi.org/10.5281/zenodo.4745814 |
|
persistent identifier |
https://treatment.plazi.org/id/03A09A04-A737-FF9D-6EDC-FCDB2810FBD8 |
|
treatment provided by |
Plazi |
|
scientific name |
Blattochaeta peterhlavaci D. Čeplík, Lakota & J. Čeplík |
| status |
sp. nov. |
Blattochaeta peterhlavaci D. Čeplík, Lakota & J. Čeplík View in CoL , sp. nov.
( Figs 5–10, 12a–12b View FIGURES 2–5 View FIGURES 6–9 View FIGURES 10, 12 )
Material studied. Holotype, ♂: MONTENEGRO, two labels: „ Montenegro , Rumija Mts., Phoenix pit, 06.08.2005, R. Mlejnek lgt.“ (white printed); „ HOLOTYPE Blattochaeta peterhlavaci sp. nov., D. Čeplík, Lakota & J. Čeplík det., 2021“ (red printed), ( CNHM) . Paratypes: 11 ♂♂, 4 ♀♀, 41 specimens sex not determined, same data as for the holotype ; 7 ♂♂, 5 ♀♀: „Montenegro, Rumija Mts., White Virgin pit, 10.08.2005; 15 ♂♂, 9 ♀♀, 37 specimens sex not determined: „ Montenegro , Rumija Mts., Ice Virgin pit, 19.05– 01.08.2005, 07.08.2005 – 27.05.2006 traps ; 1 ♂, labelled: „ Montenegro , Rumija Mts., Little Virgin pit, 05.08.2005; all R. Mlejnek lgt.“ ( CDC, CJL, CPH, CJC) . Paratypes are labelled with red printed labels „ PARATYPE Blattochaeta peterhlavaci sp. nov., D. Čeplík, Lakota & J. Čeplík det., 2021“ .
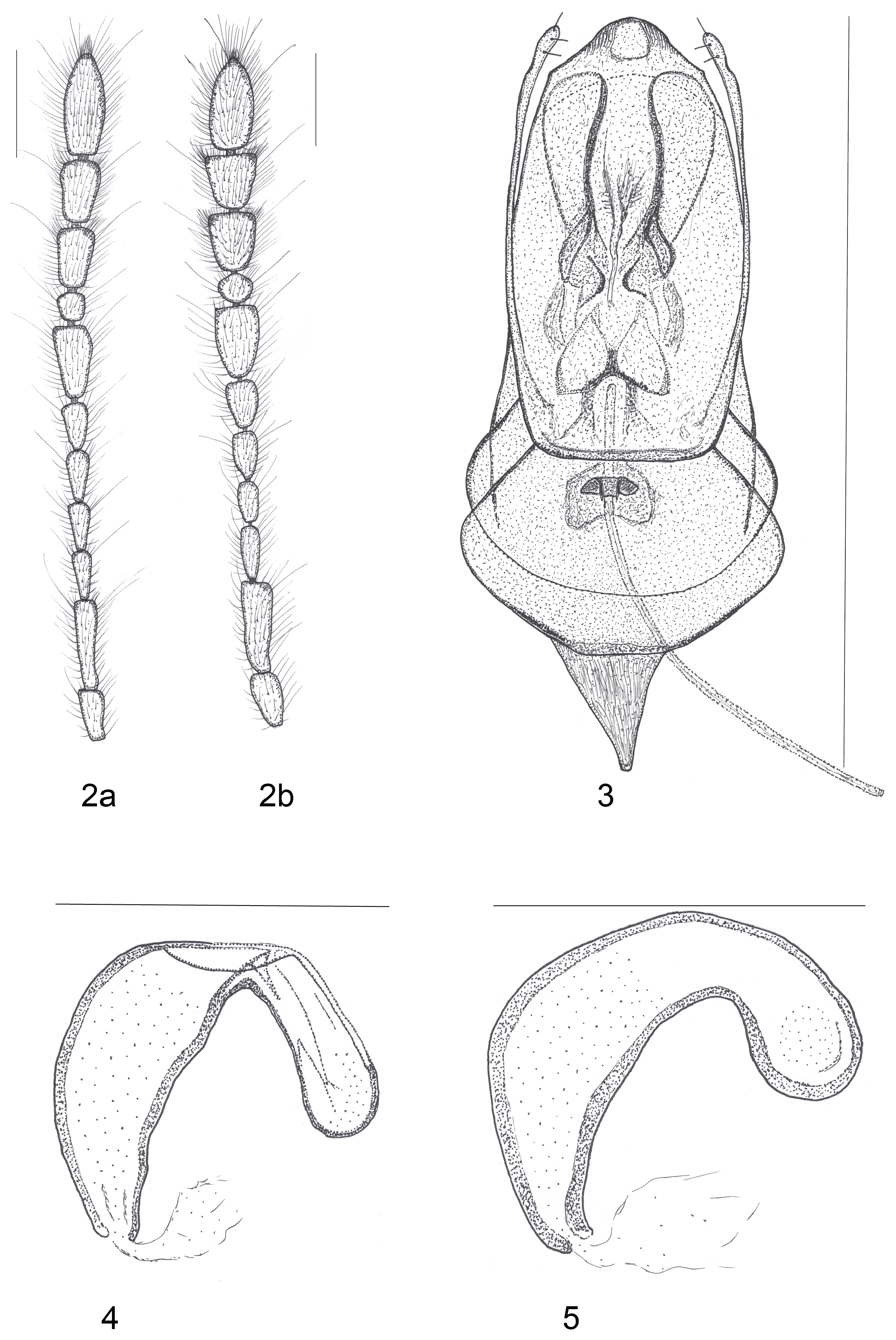
Description. Body medium–sized, bathyscioid, elliptically elongate, convex, weakly pigmented, with fine pubescence and punctures, colour from yellowish–brown to reddish–brown. Body length 4.45–4.67 mm in males, 4.50–4.85 mm in females. L: 3.83–4.22 mm. Head ( Fig. 10 View FIGURES 10, 12 ) approximately as long as wide or slightly longer than wide (due to the measuring technique of the head capable of being retracted), HL: 0.74 mm, HW: 0.74 mm, eyes completely absent, with posterior neck region and genae more or less glabrous. Transverse occipital crest present but weakly–defined. Clypeus subhexagonal, clypeofrontal suture distinct, frontoclypeus densely setose, labrum densely setose. Mandibles short, robust, with proximal and subapical incisors and one or two small teeth between. Maxillary ultimate palpomere conical, apically pointed, slender and much shorter than the thick penultimate palpomere. Antennae (AL: 2.00– 2.28 mm), finely pubescent ( Figs 8a–8b View FIGURES 6–9 ), inserted in the middle of the median third of the head. Pedicel approximately 1.60 times longer than scape. Antennomere 8 slightly longer than wide (L/W: 1.22 in males, 1.09 in females). Relative length of antennomeres (male): (1) 0.17, (2) 0.25, (3) 0.18, (4) 0.15, (5) 0.18, (6) 0.13, (7) 0.27, (8) 0.11, (9) 0.23, (10) 0.24 and (11) 0.37. Relative width of antennomeres (male): (1) 0.08, (2) 0.08, (3) 0.07, (4) 0.07, (5) 0.08, (6) 0.08, (7) 0.13, (8) 0.09, (9) 0.14, (10) 0.14 and (11) 0.13. Antennomere length/width ratios (male): (1) 2.12, (2) 3.12, (3) 2.57, (4) 2.14, (5) 2.25, (6) 1.62, (7) 2.07, (8) 1.22, (9) 1.64, (10) 1.71 and (11) 2.84. Relative length of antennomeres (female): (1) 0.17, (2) 0.29, (3) 0.18, (4) 0.15, (5) 0.18, (6) 0.14, (7) 0.26, (8) 0.12, (9) 0.20, (10) 0.20 and (11) 0.33. Relative width of antennomeres (female): (1) 0.09, (2) 0.10, (3) 0.08, (4) 0.08, (5) 0.09, (6) 0.10, (7) 0.12, (8) 0.11, (9) 0.13, (10) 0.13 and (11) 0.14. Antennomere length/width ratios (female): (1) 1.88, (2) 2.90, (3) 2.25, (4) 1.87, (5) 2.00, (6) 1.40, (7) 2.16, (8) 1.09, (9) 1.53, (10) 1.53 and (11) 2.35.
Pronotum ( Fig. 6 View FIGURES 6–9 ) trapezoidal, strongly convex, wider than long, widest at base (PL/PW: 0.52–0.60), lateral edges evenly rounded, at posterior margin widest, slightly protruding backwards, narrower than elytra. PL: 1.03–1.20 mm. PW: 1.96–2.00 mm. Dorsal surface strongly pubescent. Prosternal ventral surface sparsely pubescent.
Elytra ( Fig. 7 View FIGURES 6–9 ) elongately oval, pubescent, with maximum width at mid–length, longitudinal parasutural striae absent. EL: 2.80–3.02 mm. EW: 2.11–2.22 mm. EL/EW: 1.32–1.43. Punctation very dense, consists approximately of 20–25 punctures across the mid–width of each elytron, weak punctuation and pubescence on apical part. Scutellum widely triangular and pubescent.
Venter. Mesoventral and metaventral surface sparsely pubescent. Mesocoxal cavities separated, mesoventral process well–developed. Metacoxae separated by bifid posterior metaventral process. Mesoventral carina ( Figs 12a–12b View FIGURES 10, 12 ) developed, prolonged posteriorly, not extended over metaventrite, with backwardly oriented setae. Sternites pubescent with short, fine setae, with reticulate and leathery aspect.
Protarsi with four slender, undilated segments in both sexes, first protarsomere slightly longer than second and third, fourth longest. Mesotarsi and metatarsi pentamerous in both sexes. Claws wide and well–developed, empodium with one bifurcate seta. Protibiae armed with spines, one very distinct external subapical spine, internal subapical side with one shorter and one longer subapical multi–toothed spurs. Mesotibiae and metatibiae armed with lateral spines, with apical crown of spines (two external spines distinctly longer) of unequal length ( Gnaspini et al. 2020) and two internal apical multi–toothed spurs.
Genitalia. Aedeagus ( Fig. 9 View FIGURES 6–9 ) robust, elongate, sclerotised. Inner sac of median lobe elongate and tubular with developed chitinized structures along its length. Aedeagus 1.34–1.37 mm long, median lobe maximum width 0.44 mm, basal lamina maximum width 0.49 mm. Median lobe from dorsal aspect sub–parallel, then gradually narrower and distinctly narrowed towards pointed apex which is beak–shaped and longer than parameres. Pointed, more or less sinuate and more or less curved beak–shaped apex of the median lobe is well–visible from lateral aspect. Parameres elongate, thin, arcuate, curved inward apically, shorter than median lobe, curved before apex club which bears one apical and two lateral short setae. Basal lamina of the median lobe with large posterior expansion. Inner sac of aedeagus (endophallus) consists of apical, medial and basal sections: comparatively long and weakly sclerotized apical reinforcement bands, more sclerotized on its curved inner margins, medially positioned curved sclerotized structures, feather–like structures, connection nodules and basal phanera; ejaculatory duct is illustrated. Spermatheca with basal, only slightly narrow medial and apical regions ( Fig. 5 View FIGURES 2–5 ). Female ventrite VIII densely pubescent with anterior expansion stout. Each style with four setae, stylus cylindrical, with one long seta.
Sexual dimorphism:Female slightly larger than male.Antennal length 2.22–2.28 mm in males and 2.00– 2.22 mm in females. Antennomere 8, ratio L/W: 1.22 in males and 1.09 in females. Shape of mesoventral carina variable.
Intra–species morphological variability (inside the population from the type locality, between and inside populations from all recorded localities): slightly variable shape of the elytra; more or less variable shape of the mesoventral carina (visible from lateral aspect); slightly variable structures of inner sac of the median lobe; slightly variable shape of posterior expansion of the aedeagus; variable shape of more or less sinuate–more or less curved beak–shaped apex of the median lobe (visible from lateral aspect); slightly variable shape of the spermatheca.
Differential diagnosis and remarks. Blattochaeta peterhlavaci sp. nov. differs from other species by antennomere 8 slightly longer than wide (L/W: 1.22 in males, 1.09 in females) versus antennomere 8 approximately as long as wide (in male L/W: 1.10, in females L/W: 0.91, Blattochaeta brankojalzici sp. nov.), or antennomere 8 much longer than wide (L/W: 1.72 in male, Blattochaeta remyi ; L/W: 1.66 in male, Blattochaeta montenegrina ; L/W: 1.80–2.02, Blattochaeta marianii kusijanovici ; L/W: 1.50–2.50, other studied specimens of Blattochaeta marianii ssps.; L/W: approximately 2.00, Blattochaeta matchai , from Jeannel, 1924). Related available species of the genus can be separated from Blattochaeta peterhlavaci sp. nov. further as follows:
– Blattochaeta brankojalzici sp. nov. by: sparse punctation on elytra, which is approximately 18–20 punctures across the mid–width of each elytron (versus dense, approximately 20–25 punctures in Blattochaeta peterhlavaci sp. nov.); basal lamina maximum width 0.59 mm (versus basal lamina maximum width 0.49 in Blattochaeta peterhlavaci sp. nov.); spermatheca with distinctly narrow medial region (versus spermatheca with only slightly narrow medial region in Blattochaeta peterhlavaci sp. nov.); other differences in aedeagi and spermathecae of both species are shown in the accompanying figures.
– Blattochaeta remyi by: total body length approximately 4.30 mm in male, length approximately 3.80 mm in male (versus total body length 4.45–4.85 mm, length 3.83–4.22 mm in new species); EW: 1.94 mm in male (versus EW: 2.11–2.22 mm in new species); sparse punctation on elytra, which is approximately 18–20 punctures across the mid–width of each elytron (versus dense, approximately 20–25 punctures in new species); antennal length 2.40 mm in male (versus antennal length 2.00– 2.28 mm in new species); aedeagus length 1.10 mm (versus aedeagus length 1.34–1.37 mm in new species); basal lamina maximum width 0.41 mm (versus basal lamina maximum width 0.49 mm in new species); median lobe maximum width 0.34 mm (versus median lobe maximum width 0.44 mm in new species); parameres slightly longer than median lobe (versus parameres shorter than median lobe in new species); other differences on aedeagi of both species are shown in the accompanying figures.
– Blattochaeta montenegrina by: total body length approximately 5.00 mm in male, length approximately 4.30 mm in male (versus total body length 4.45–4.85 mm, length 3.83–4.22 mm in new species); EW: 2.34 in male (versus EW: 2.11–2.22 mm in new species); antennal length 2.85 mm in male (versus antennal length 2.00– 2.28 mm in new species); sparse punctation on elytra, approximately 15 punctures across the mid–width of each elytron (versus denser punctation on elytra, approximately 20–25 punctures in new species); basal lamina maximum width 0.58 mm (versus basal lamina maximum width 0.49 mm in new species); other differences on aedeagi of both species are shown in the accompanying figures.
– Blattochaeta marianii ssps. and B. matchai by: sparse punctation on elytra, approximately 18–20 punctures across the mid–width of each elytron (versus denser punctation on elytra, approximately 20–25 punctures in new species); antennal length 2.50–3.20 mm (versus antennal length 2.00– 2.28 mm in new species); total body length 4.85–5.80 mm, length 4.20–5.00 mm (versus total body length 4.45–4.85 mm, length 3.83–4.22 mm in new species); EW: 2.30–2.70 mm (versus EW: 2.11–2.22 mm in new species).
Etymology. Dedicated to our dear friend Peter Hlaváč (Prague, Czech Republic), a specialist for selected groups of Staphylinidae and Curculionidae and member of Biospeleologica Slovaca.
Distribution and data. The new species is known from its type locality, the Phoenix pit (– 70 m), at the elevation of approximately 1100 m a.s.l., and from the nearby White Virgin pit, Ice Virgin pit and Little Virgin pit in Rumija Mts., southeastern Montenegro. The Phoenix pit was described in Lohaj et al. 2016. The new species probably inhabits more speleological localities situated in Rumija Mts. Here are stated translated data from Pretner (1977: 127, 139, 15): “a small female of Blattochaeta , perhaps new species, from a little pit, snow cave near Miligrepa, Mali Mikulići, Rumija Mts, 05.07.1960, Dušan Gavrilovićˮ. This specimen was not studied for this work.
Associated subterranean Coleoptera from the type locality: Adriaphaenops rumijaensis Lohaj et al. 2016 , Neotrechus suturalis ssp. Schaufuss, 1864, Laemostenus (Antisphodrus) cavicola ssp. Schaum, 1858, Anthroherpon matulici Reitter, 1903 , and Anthroherpon taxi ssp. Müller, 1913, from Lohaj et al. 2016.


| R |
Departamento de Geologia, Universidad de Chile |
| CNHM |
Cincinnati Museum of Natural History |
| CDC |
Changdu Institute for Drug Control |
| CPH |
University of the Pacific |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |