
Varicus, ROBINS
|
publication ID |
https://doi.org/ 10.1111/zoj.12394 |
|
publication LSID |
lsid:zoobank.org:pub:E952647E-1571-4A14-8BD4-54D1746760D0 |
|
persistent identifier |
https://treatment.plazi.org/id/03A0C25D-BB79-FFB1-B7C2-FC40FD59E828 |
|
treatment provided by |
Marcus |
|
scientific name |
Varicus |
| status |
|
VARICUS ROBINS View in CoL View at ENA & B OHLKE €, 1961
TYPE SPECIES: VARICUS BUCCA ROBINS & B OHLKE €, 1961 (P. 47, FIGS 1 – 3 View Figure 1 View Figure 2 View Figure 3 , BY ORIGINAL DESIGNATION)
Diagnosis
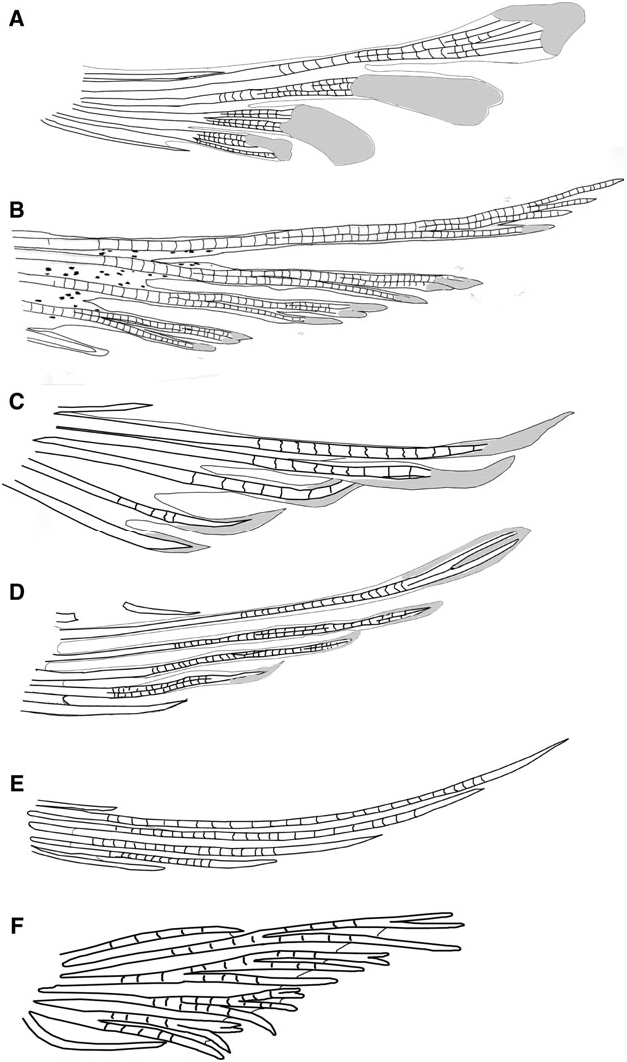
Possesses all taxonomic characters present in most members of Gobiosomatini and the Gobiosoma group (first dorsal-fin spines VII, pterygiophore insertion pattern of 3 – 221110, 27 vertebrae – 11 precaudal and 16 caudal (rarely 12 and 15), hypurals 1 and 2 fused to some extent with hypurals 3 and 4 and the terminal vertebral element, one epural); pelvic fins well separated, lacking both anterior frenum and well-developed membrane connecting innermost rays; pelvic-fin rays highly variable among species ( Fig. 4A – E View Figure 4 ), with rays 1 – 4 unbranched, branched internally and re-fused with pointed fleshy pads, or branched to tips with flat, spatulate fleshy pads, fifth pelvic-fin ray always unbranched; pelvic-fin rays generally extending posteriorly to anus and sometimes to origin of rays 1 – 4 of anal fin; body with scales in all but one species ( Varicus decorum sp. nov.) (modified basicaudal scales present in all but one species, which awaits description; see remarks below); one anal-fin pterygiophore inserted before first haemal spine (in anomalous cases where first haemal spine is on vertebra 13 rather than 12, two anal-fin pterygiophores inserted before haemal spine); papillae rows 5i and 5s connected in all but three species ( Varicus benthonis , Varicus vespa , and variable in Varicus cephalocellatus sp. nov.); cephalic lateralis canals and pores absent; second dorsal-fin rays I,8 – 10; anal-fin rays I,7 – 9 (rarely I, 6 in one species); body coloration variable, but generally with yellow spots or stripes on dorsal and caudal fins, bright-yellow spots, bands, or saddles on dorsal and lateral surface of body, and in several species, dark vertical bars or bands along side of body. The ten species in this genus are known only from the western Atlantic Ocean.
Remarks on the genus Varicus
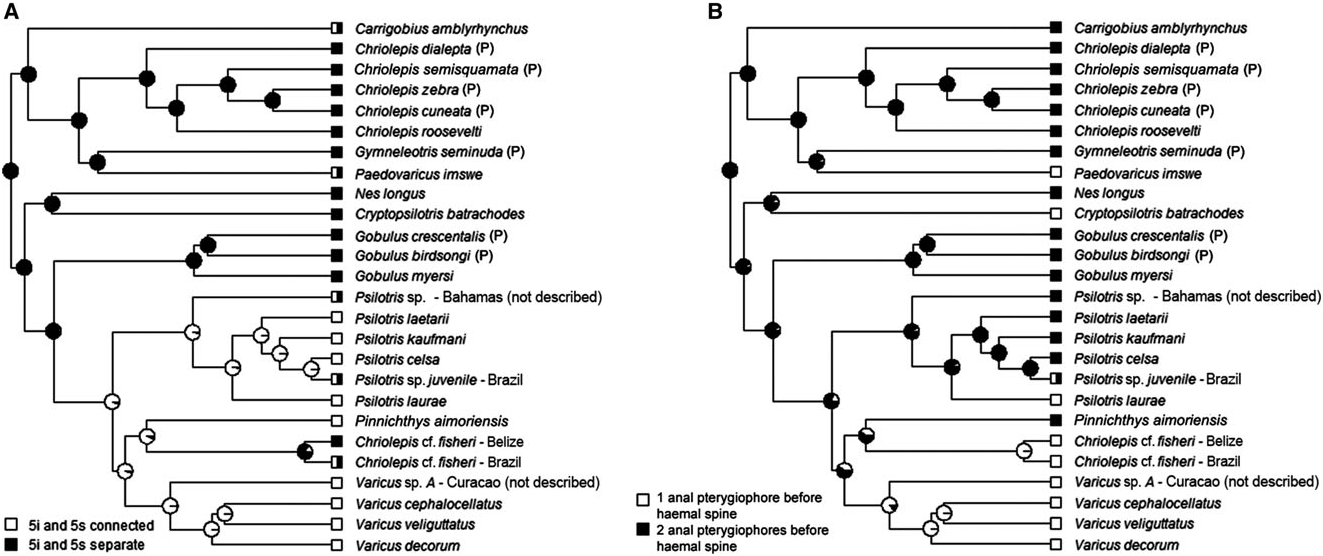
Varicus View in CoL was described by Robins & Bohlke (1961) for Varicus bucca View in CoL . Since then Varicus View in CoL has been loosely distinguished from Chriolepis View in CoL based on Varicus View in CoL having unbranched pelvic-fin rays (rays branched to the tips in Chriolepis View in CoL ), having a bilobed tongue (rounded in Chriolepis View in CoL ), and in lacking a low fleshy ridge between the innermost pelvic-fin rays (ridge present in Chriolepis View in CoL ); however, as more species from both genera were subsequently described, mostly from the Atlantic (e.g. Chriolepis vespa View in CoL , Varicus marilynae Gilmore, 1979 View in CoL , and Varicus imswe View in CoL ), the generic distinctions began to blur, and the aforementioned characters proved to be of little use in separating species into two clearly delineated groups ( Hastings & Bortone, 1981). Birdsong et al. (1988) noted that Varicus View in CoL and the Atlantic members of Chriolepis View in CoL ( Chriolepis vespa View in CoL , Chriolepis fisheri View in CoL , and Chriolepis benthonis View in CoL ) are the only species of Gobiosomatini to have one anal-fin pterygiophore inserted anterior to the first haemal spine (two in all other species), indicating a possible close relationship between the two groups and a potential distinction between the Atlantic and Pacific species of Chriolepis View in CoL ; however, this link between the pterygiophore pattern and the divide between Atlantic and Pacific species became less apparent when Hastings & Findley (2013, 2015) described two Atlantic species that have two anal-fin pterygiophores anterior to the first haemal spine, tentatively placing both in Chriolepis View in CoL .
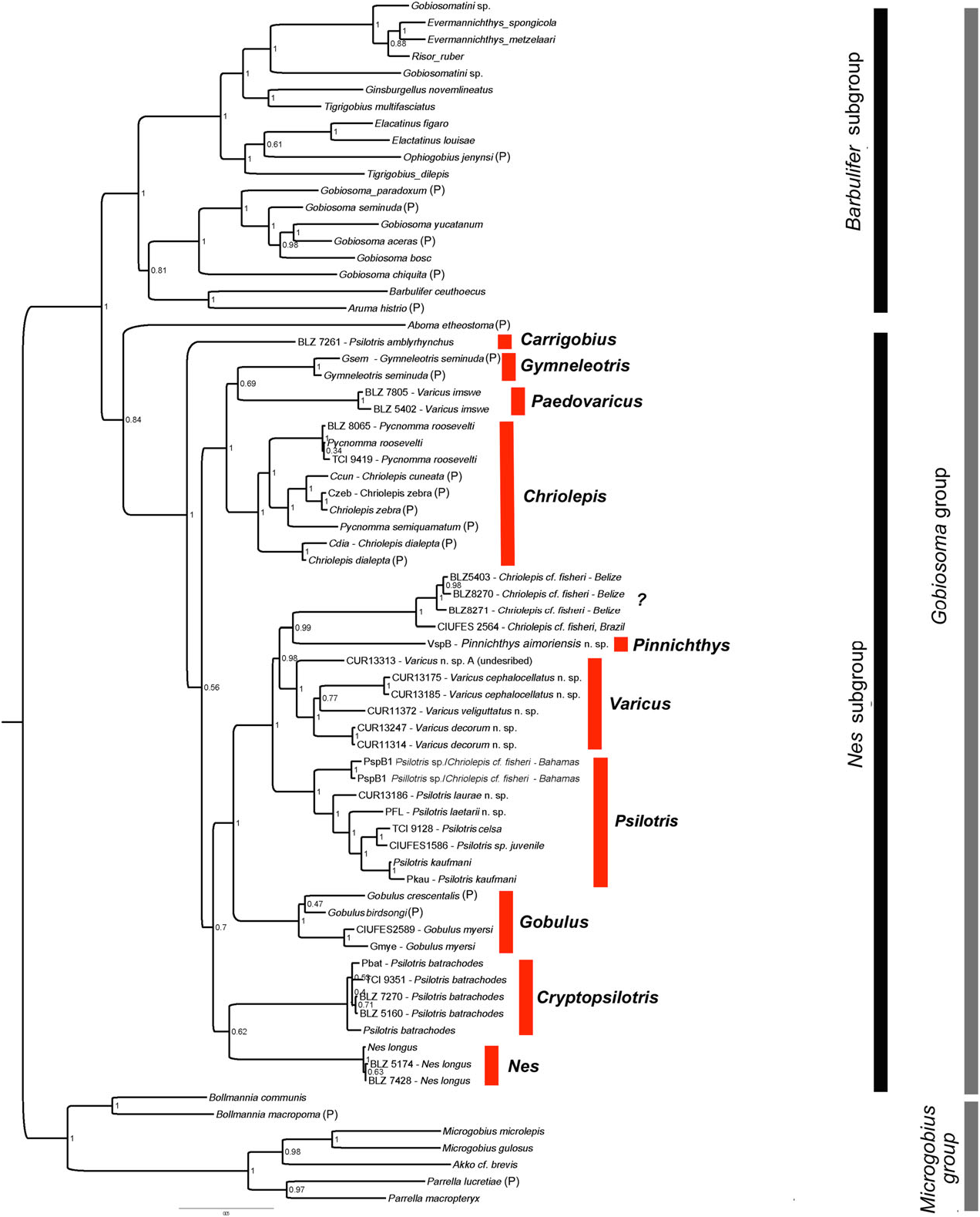
Our phylogenetic analysis includes only one named species of Varicus View in CoL ( Varicus imswe View in CoL ); however, several other deep-reef Caribbean species are included in our tree that are very similar to the type species, Varicus bucca View in CoL , and are likely to be much closer relatives to Varicus bucca View in CoL than to Varicus imswe View in CoL . The four species in our tree that are similar to Varicus bucca View in CoL are the new species Varicus cephalocellatus sp. nov., Varicus decorum sp. nov., and Varicus veliguttatus sp. nov., plus one undescribed species ( Varicus sp. A in Fig. 5 View Figure 5 ). The four species form a well-supported monophyletic group, and all have one anal-fin pterygiophore inserted before the first haemal spine, pelvic-fin rays that are either unbranched or branched internally, and pelvic-fin rays with fleshy tips. Those morphological characters are shared with Varicus bucca View in CoL and Varicus marilynae View in CoL . Varicus imswe View in CoL is distinct from these four species both genetically and morphologically, and is herein assigned to a new genus Paedovaricus View in CoL (see remarks section for Paedovaricus View in CoL above for justification). In addition to the aforementioned species in our tree, two other new species are described here without molecular data and placed in Varicus View in CoL ( Varicus adamsi sp. nov. and Varicus nigritus sp. nov.). These two also possess one anal-fin pterygiophore before the first haemal spine and pelvic-fin rays that are either unbranched or are branched but have fleshy tips. The anal-fin pterygiophore character is not entirely free of homoplasy ( Fig. 6B View Figure 6 ), but it nevertheless contains considerable phylogenetic information ( Table 1); thus, it may be of some utility in classifying species, if not by itself then in combination with other characters (e.g. pelvic-fin branching pattern, fin-ray counts, papillae pattern, etc.). In addition, most species in this group have papillae rows 5i and 5s connected to form a single continuous row (variable in Varicus cephalocellatus sp. nov., separate in Varicus vespa and Varicus benthonis , see paragraph below). This character is apparently derived in the Nes View in CoL subgroup and is shared among several lineages of Atlantic species. Another important characteristic of the species in our Varicus View in CoL clade is that, like Varicus bucca View in CoL and Varicus marilynae View in CoL , all species occur on continental or insular slopes at or below 60 – 100 m depth. Collectively, despite lacking a single unambiguous morphologial synapomorphy, we recognize the five new species described here plus Varicus marilynae View in CoL and Varicus bucca View in CoL as belonging to the genus Varicus View in CoL based on the combined molecular, morphological, and ecological information presented above.
Of the five Atlantic species of Chriolepis , two are now referred to the new deep-reef genus Pinnichthys based on the presence of two anal-fin pterygiophores before the first haemal spine (versus one in Varicus ), a high number of lateral scale rows (≥30 vs. ≤ 27 in Varicus ), and more anal-fin rays (I,10 vs. I,9 or fewer in Varicus ). The status of Chriolepis fisheri remains unclear (see remarks section for Chriolepis above). The two remaining species, Chriolepis vespa and Chriolepis benthonis , are considered here to belong to Varicus rather than Pinnichthys or Chriolepis based on the presence of one anal-fin pterygiophore inserted before the first haemal spine, a low number of lateral scales (≤12), low anal-fin ray counts (I,8 or fewer), and the fact that both are deep-water species. This assignment is tentative, however, as papillae rows 5i and 5s are separate in both species (nearly connected in Varicus vespa ), and both species possess highly branched pelvic-fin rays (some rays in Varicus vespa and Varicus benthonis with minute fleshy tips at the end of some branches, Fig. 4B View Figure 4 ). Varicus ben- thonis is known only from the holotype, which is in poor condition. Additional fresh specimens and molecular data from these species may ultimately prove helpful in verifying their taxonomic placement. Lastly, Varicus overlaps with Psilotris to some extent for virtually every morphological feature examined here. Details on morphologically differentiating Psilotris and Varicus are given in the remarks section for Psilotris .
Remarks on the type species Varicus bucca and unidentified Varicus specimens
With the recent accumulation of many new specimens of Varicus and the discovery of multiple new species in this genus, we have re-examined the type series of Varicus bucca and come to several conclusions. First, the type series probably contains multiple species, given the wide range of second dorsal-fin and anal-fin counts present among specimens. Furthermore, several of the paratypes have broken pelvic and pectoral rays, and the pigmentation for all specimens has faded, making identification difficult. For these reasons we consider only specimens with I,9 second dorsal-fin rays and I,7 – 8 anal-fin rays to be Varicus bucca , and some of those only tentatively so. We provide an annotated list of Varicus specimens that are confidently identified as Varicus bucca as well as those that are of questionable identity, including some paratypes of Varicus bucca ( Table S1).
VARICUS ADAMSI GILMORE, VAN TASSELL & TORNABENE SP. NOV. TWILIGHT GOBY
FIGS 23 – 26 View Figure 23 View Figure 24 View Figure 25 View Figure 26
Holotype
USNM 427226 About USNM , 60 About USNM mm SL, male, Johnson Sea Link I Dive JLS-I-2027, 23.957833N – 74.57066W, Sand Point , San Salvador, Bahamas, 435.1 m depth, 4 May 1987, R. G. Gilmore & M. Adams. GoogleMaps
Paratype
USNM 427225 About USNM , 44.5 mm SL, female, Johnson Sea Link I Dive JLS-I-2022, 24.050833N – 74.53933W, Cockburn Town , Riding Rock, San Salvador, Bahamas, 280 m depth, 2 May 1982 GoogleMaps , R. G. Gilmore & M. Adams ; USNM 220985 About USNM , 32.5 mm SL male , R / V Oregon Cruise 94, Station 5021, 11.3533N — 60.645W, north of Tobago, tumbler dredge, Trinidad and Tobago, 165 – 183 m depth, 21 September 1964 GoogleMaps .
Diagnosis
Side of body typically with ctenoid scales from caudal peduncle to pectoral-fin base, ctenii reduced or lack- ing anteriorly; six or seven very enlarged ctenoid scales on caudal peduncle; two modified basicaudal scales with enlarged ctenii on dorsal and ventral margins of caudal-fin base; first dorsal-fin spines VII, no elongate spines; second dorsal-fin rays I,9; analfin rays I,8; pectoral fin extending to third element of second dorsal fin; pelvic fins well separated, fifth ray one-quarter length of fifth ray, rays 1 – 4 branched several times with spatulate, fleshy tips; papillae rows 5s and 5i connected as a continuous transverse row; interorbital papillae row pc’ with two papillae; no cephalic lateralis pores on head or preopercle; anterior nostril with dorsal bilobed flap, posterior with simple triangular flap; one anal-fin pterygiophore inserted anterior to haemal arch; pectoral, spinous dorsal and caudal fins with brilliant white pigment adjacent to black fin margins, body white with seven conspicuous yellow bars situated under eye, on cheek, behind opercular margin, below middle of dorsal fin, below origin of second dorsal fin, below middle of second dorsal fin, and on posterior margin of caudal peduncle.
Description
Morphometric data are presented in Table 3.
Median and paired fins: First dorsal-fin spines VII (3), spines 2 – 4 extending slightly from membrane, none notably elongate or filamentous; second dorsal fin I, 9(3); anal fin I,8*(2), I,7(1); pectoral-fin rays 18 (3), rays 13 and 14 longer than others, extending to vertical through third ray of second dorsal fin; pelvic fin I,5(3); pelvic fins well separated, lacking anterior frenum and membrane connecting bases of innermost rays; fourth pelvic-fin ray longest extending to anus, pelvic-fin rays 1 – 4 branched several times, tips of branches united with fleshy spatulate pads ( Fig. 4A View Figure 4 ), fifth pelvic-fin ray unbranched, short, one-quarter length of fourth pelvic-fin ray; caudal fin lanceolate in male, truncate in female; branched caudal-fin rays 14(3); segmented caudal-fin rays 17(3).
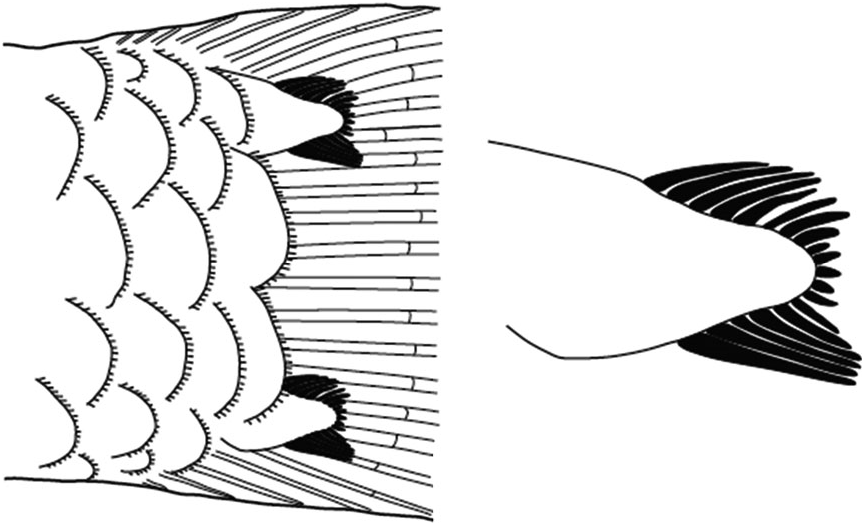
Scales: Trunk of body with ctenoid scales, length of ctenii reduced anteriorly; six or seven very enlarged ctenoid scales on caudal peduncle; 21 – 24 lateral scale rows extending to beneath pectoral fin in holotype (USNM 427226, male), male paratype (USNM 220985) with 23 lateral scale rows extending to beneath pectoral fin, female paratype (USNM 427225) with only enlarged scales on caudal peduncle, remainder of body abraded; two enlarged modified basicaudal scales with enlarged ctenii present at dorsal and ventral margins of caudal-fin base.
Head: Jaw extending posteriorly to a vertical through either the anterior margin of eye or the anterior margin of pupil; anterior nare as a short tube with dorsal bilobed flap; posterior nare an opening with simple triangular flap; no cephalic lateralis pores on head or preopercle; eyes 30.6 – 41.8% HL, dorsolateral, extending above head profile; interorbital narrow 4.3 – 5.1% HL; snout profile steep; operculum opening extending only length of pectoral-fin base; teeth (observed from USNM 220985) in both jaws consisting of multiple irregular rows of small conical teeth, no enlarged canines.
Genitalia: Male genitalia pointed and triangular; female as bulbous pore; no melanophores present on either sex.
Colour in life ( Figs 23 View Figure 23 and 24 View Figure 24 ): Background of body pale white; trunk with four narrow bright yellow bars that traverse the dorsal midline, diameter greater than pupil but less than eye; first trunk bar located below middle of first dorsal fin; second trunk bar below origin of second dorsal fin; third trunk bar below middle of second dorsal fin; last trunk bar over posterior end of caudal peduncle; head with three narrow vertical yellow bars; first vertical bar on head directly below eye; second vertical bar on head extending ventrally from anterior nape onto margin of preopercle; third vertical bar on head extending ventrally from posterior nape onto upper opercular margin; iris of eye golden yellow; tip of snout yellow; opercle pale pink.
Coloration of fins in the male holotype differs from female paratype. The following description is based on the male: first dorsal fin covered with white pigment except on spines, with yellow – orange pigment on basal third of spines opposite the first vertical yellow body bar, distal margin black; second dorsal with faint yellow coloration on basal portion of rays at location of vertical yellow body bars; black distal margin; caudal fin white; fin margin with black pigment; anal fin with black on lower third of fin; rays adjacent to yellow body bars with yellow pigment; distal third of pectoral fin with black pigment, remainder of fin with white pigment. Female pectoral fins and anal fin lack black pigment and dorsal and caudal fins have much reduced black pigment compared with the male.
Colour in preservation ( Fig. 25 View Figure 25 ): Background colour tan with faint vertical melanic bars at locations where yellow bars were observed on head and trunk of fresh specimens; dusky distal margins of dorsal, anal, and caudal fins retaining pigment in some specimens, faded in others.
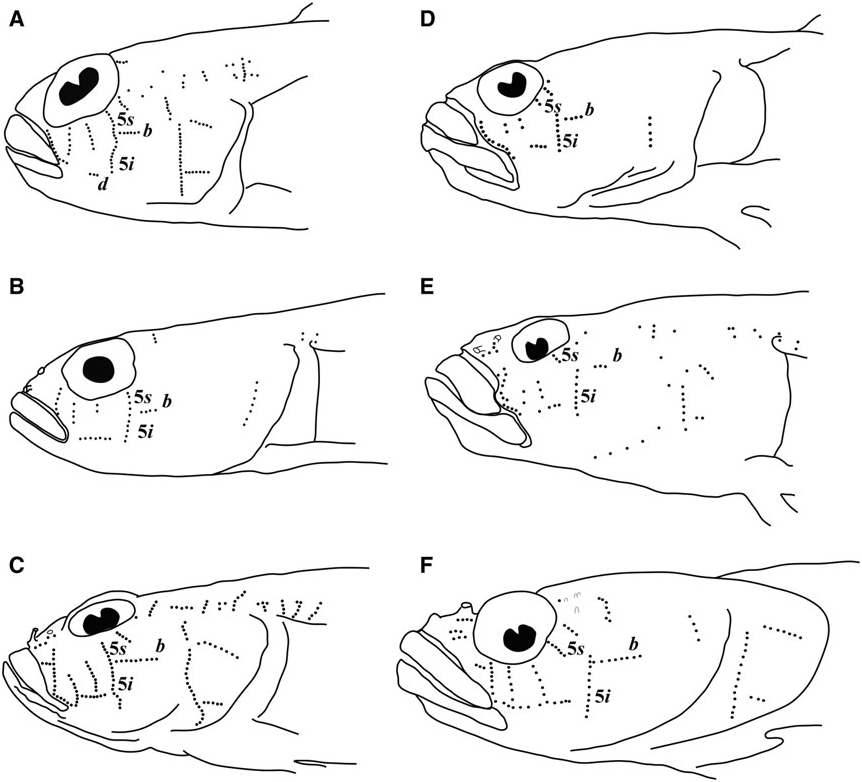
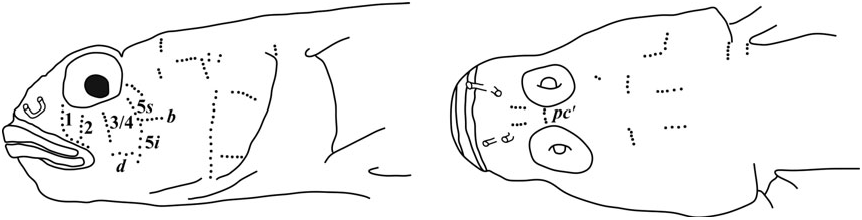
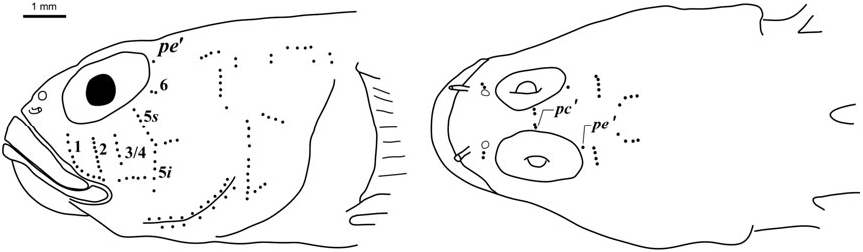
Sensory papillae ( Fig. 26 View Figure 26 ): A transverse pattern with rows 1, 2, 3/ 4, 5s /5i, and 6 present; row 5s/5i joined forming a single row and extending below the level of row d; row b short with four or five papillae, ending anteriorly at row 5s/5i; interorbital row pc’ with two papillae.
Vertebral skeleton: Dorsal pterygiophore formula 3 – 221110; one anal-fin pterygiophore inserted anterior to first haemal arch; hypurals 1 and 2 fused with hypurals 3 and 4 along one-quarter of their length; 27 vertebrae – 11 precaudal and 16 caudal.
Habitat: This is the deepest occurring goby of the genus Varicus , and possibly the deepest occurring goby in the world, and has been captured at depths of 280 and 435.1 m during manned submarine dives, JSL – II, 2022 and 2027 off the western shore of San Salvador , Bahamas. The substrate at the type locality principally consists of limestone outcroppings and boulders with an occasional thin layer of calcareous oolite sands and Halimeda spp. algal rubble (see in situ photo of the holotype at 435 m off Sandy Point, San Salvador; Fig. 24 View Figure 24 ). Bare sedimentary rock surface and boulders predominate in this region at depths of 250 – 500 m.
Distribution: Known only from the western shore of San Salvador, Bahamas and from Tobago.
Etymology: Named for the late famed research submersible pilot, Mr. Michael Adams, who painstakingly captured both Bahamas specimens during a 30 – 45 min chase using a 26 ton submarine (JSL – II) in simultaneous multiple thrust, multidirectional mode to allow this description. Mr. Adams was one of the five original research submersible pilots within the United States.
Comparisons: Varicus adamsi sp. nov. is distinctive in that it is the only species of Varicus to have highly branched pelvic-fin rays 1 – 4 that terminate in flattened, spatulate fleshy tips ( Fig. 4A View Figure 4 ). The I,9 count for second dorsal-fin rays further distinguishes Varicus adamsi sp. nov. from Varicus cephalocellatus sp. nov. (I,10) and from Varicus marilynae , Varicus benthonis and Varicus veliguttatus sp. nov. (all with I,8). Varicus adamsi sp. nov. is further distinguished from Varicus nigritus sp. nov. in lacking prominent black wide bands along the body. The presence of body scales easily distinguish Varicus adamsi sp. nov. from Varicus decorum sp. nov. Varicus adamsi sp. nov. is likely most closely related to Varicus vespa . The two species have similar meristic counts and nearly identical coloration ( Figs 23 View Figure 23 , 24 View Figure 24 , and Fig. S4D View Figure 4 ). Both Varicus adamsi sp. nov. and Varicus vespa have branched pelvic rays, although rays in Varicus vespa never have the distinct spatulate tips. Varicus vespa is a relatively small species (types <30 mm SL) known only from the Gulf of Mexico where it occurs over mud, whereas Varicus adamsi sp. nov. is larger (reaching 61 mm SL) and is known from the Bahamas and Tobago where it occurs over hard calcareous substrate. The differences in distribution, maximum size, habitat association, and pelvic-fin morphology support that the two represent distinct species.
VARICUS CEPHALOCELLATUS N. SP., GILMORE, VAN TASSELL, AND BALDWIN
FIGS 27 – 30 OCELLATED View Figure 27 View Figure 28 View Figure 29 View Figure 30 SPLITFIN GOBY
Holotype
USNM 427232 About USNM , 28.2 mm SL, female, Paynes Bay , Barbados, West Indies, 13.162N — 59.658W, Johnson Sea Link II Dive JLS-II 1755, 159 m depth, 1 May 1989, C.L. Smith & C. Caddigan. GoogleMaps
Paratypes
USNM 426788 About USNM , 30.4 mm SL, female, Bonaire City Dock , Kralendijk, Bonaire, Lesser Antilles, 12.15N, — 68.2829W, tissue CUR13185, 114 – 137 m depth, 5 May 2013 GoogleMaps , Dive 2 CURASUB submersible, B. Van Bebber, A. Schrier, C. Baldwin , T. Christiaan ; USNM 426736 About USNM , 39.8 mm SL, female, Bonaire City Dock , Kralendijk, Bonaire, Lesser Antilles, 12.15N, — 68.2829W, tissue CUR13175, 122 – 140 m depth, 5 May 2013 GoogleMaps , Dive 1, CURASUB submersible, B. Van Bebber, A. Schrier, C. Baldwin , & T. Christiaan ; USNM 427227 About USNM , 37.2 About USNM / 37.08 mm SL, 1 male and 1 female, Johnson Sea Link II Dive JSL-II 1772, 13.34N — 61.2266W, Larikai Point, St. Vincent, 144 m depth, 9 May 1989 GoogleMaps , R. G. Gilmore & C. Caddigan .
Diagnosis
Side of body with 12 – 23 ctenoid scales extending anteriorly to between vertical under the middle of the second dorsal fin, to just posterior of pectoral-fin base, scales becoming smaller with reduced ctenii anteriorly; modified basicaudal scales present; first dorsal-fin spines VII; second dorsal-fin rays I,10; anal-fin rays I,9; pectoral fin with 19 – 20 rays, (23.9 – 30.6% SL), with rays 13 – 16 greatly extended, reaching base of anal-fin rays 1 – 3, giving fin asymmetrical appearance; fleshy exertions on tips of spinous dorsal, first six rays of second dorsal; pelvic fins well separated lacking anterior frenum and membrane connecting innermost rays, 5th pelvic-fin ray 1/4 – 1/5 length of 4th ray, all rays unbranched or branched internally and re-fused, with flattened fleshy tips, longest ray extending posteriorly to anal-fin origin or reaching origin of 2nd anal-fin ray; sensory papillae rows 5 s and 5 i connected as a continuous transverse row, separated by a space of 1 – 2 papillae in two specimens; interorbital papillae pc ’ and pe ’ present; no cephalic lateralis pores on head or preopercle; one anal-fin pterygiophore inserted anterior to haemal arch; series of 3 – 5 conspicuous yellow ocelli on head extending from mouth to nape over dense field of numerous red, brown and gold chromatophores.
Description
Morphometric data are presented in Table 3.
Median and paired fins: First dorsal-fin spines VII (5), spines 2 – 4 nearly equal in length, no elongate spines, fleshy exertions present on holotype and two paratypes; second dorsal fin I,10(5), spine and rays of second dorsal fin with 1 – 4 with exerted fleshy tips (holotype and two paratypes), soft rays 1 – 4 undivided or with minor terminal bifurcation, remainder of second dorsal-fin rays bifurcated for 30 – 50% of element length; anal fin I,9(5); pectoral-fin rays 20*(4), 19(1), rays 13 – 16 greatly extended beyond remainder of fin reaching vertical through anal-fin rays 1 to 3; pelvic-fin rays I,5(5); pelvic fins well separated, lacking both anterior frenum and membrane connecting bases of innermost rays; 4th pelvic-fin ray longest extending to anal-fin origin or 2nd anal-fin ray, reaching origin of 2nd anal-fin ray in one specimen; 5th pelvic-fin ray 1/4 – 1/5 length of 4th ray; all pelvic-fin rays unbranched, or branched internally but re-fused at tips, all tips generally with single fleshy pads; caudal fin rounded to truncate or slightly emarginate; branched caudal rays 14*(4) 15







| R |
Departamento de Geologia, Universidad de Chile |
| V |
Royal British Columbia Museum - Herbarium |
| T |
Tavera, Department of Geology and Geophysics |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Varicus
| Tornabene, Luke, Van Tassell, James L., Gilmore, Richard G., Robertson, David Ross, Young, Forrest & Baldwin, Carole C. 2016 |
Chriolepis fisheri
| Tornabene & Van Tassell & Gilmore & Robertson & Young & Baldwin 2016 |
Varicus cephalocellatus
| Tornabene & Van Tassell & Gilmore & Robertson & Young & Baldwin 2016 |
Varicus decorum
| Tornabene & Van Tassell & Gilmore & Robertson & Young & Baldwin 2016 |
Varicus veliguttatus
| Tornabene & Van Tassell & Gilmore & Robertson & Young & Baldwin 2016 |
Paedovaricus
| VAN TASSELL, TORNABENE & GILMORE 2016 |
Paedovaricus
| VAN TASSELL, TORNABENE & GILMORE 2016 |
Varicus adamsi
| Tornabene & Van Tassell & Gilmore & Robertson & Young & Baldwin 2016 |
Varicus nigritus
| Tornabene & Van Tassell & Gilmore & Robertson & Young & Baldwin 2016 |
Varicus cephalocellatus
| Tornabene & Van Tassell & Gilmore & Robertson & Young & Baldwin 2016 |
Chriolepis vespa
| Hastings & Bortone 1981 |
Varicus imswe
| Greenfield 1981 |
Chriolepis vespa
| Hastings & Bortone 1981 |
Varicus imswe
| Greenfield 1981 |
Varicus imswe
| Greenfield 1981 |
Varicus imswe
| Greenfield 1981 |
Varicus vespa
| Hastings & Bortone 1981 |
Varicus marilynae
| Gilmore 1979 |
Varicus marilynae
| Gilmore 1979 |
Varicus marilynae
| Gilmore 1979 |
Varicus marilynae
| Gilmore 1979 |
Chriolepis benthonis
| Ginsburg 1953 |
Nes
| GINSBURG 1933 |
Chriolepis
| GILBERT 1892 |
Chriolepis
| GILBERT 1892 |
Chriolepis
| GILBERT 1892 |
Chriolepis
| GILBERT 1892 |
Chriolepis
| GILBERT 1892 |
Chriolepis
| GILBERT 1892 |
Chriolepis
| GILBERT 1892 |