
Integripelta, Gordon & Mawatari & Kajihara, 2002
|
publication ID |
https://doi.org/ 10.1046/j.1096-3642.2002.00020.x |
|
persistent identifier |
https://treatment.plazi.org/id/03A1878A-FFB8-FF85-FC0F-FC22FE12E718 |
|
treatment provided by |
Carolina |
|
scientific name |
Integripelta |
| status |
gen. nov. |
INTEGRIPELTA View in CoL GEN. NOV.
Diagnosis
Colony encrusting, multiserial. Autozooids with smooth gymnocystal frontal shield lacking foramina. Zooidal orifices with proximolateral indentations, and shallow excavations in the adjacent gymnocyst; oral rim lacking median suboral suture. No spines or avicularia. Maternal orifice usually slightly larger than autozooidal orifice, or obviously so; brooding internal, with a distal ovicell-like kenozooid with a central perforation. Ancestrula with membranous frontal wall only, no spines. Interzooidal communications via uniporous mural septula.
Type species: Lepralia bilabiata Hincks 1884 .
Etymology
From integer (L.) whole, and pelta (f. L.) a small shield.
Remarks
A new genus is established here for encrusting eurystomellids with imperforate frontal shields and communicating via uniporous mural septula instead of basal pore-chambers. In the diagnosis, the ancestrula is described as having a membranous frontal wall only. This is based on the description of I. bilabiata by Soule et al. (1995), who described it as resembling the adult zooid “but with cuticular frontal wall”; there are no spines (D. Soule, pers. comm. 2001). This accords with the form of the ancestrula in Eurystomella , which has no frontal shield and the distal rim and operculum are like those of later zooids. The ancestrula has not yet been encountered in any of the new species described below. Integripelta bilabiata was fully described and illustrated by SEM by Soule et al. (1995).
INTEGRIPELTA NOVELLA SP. NOV.
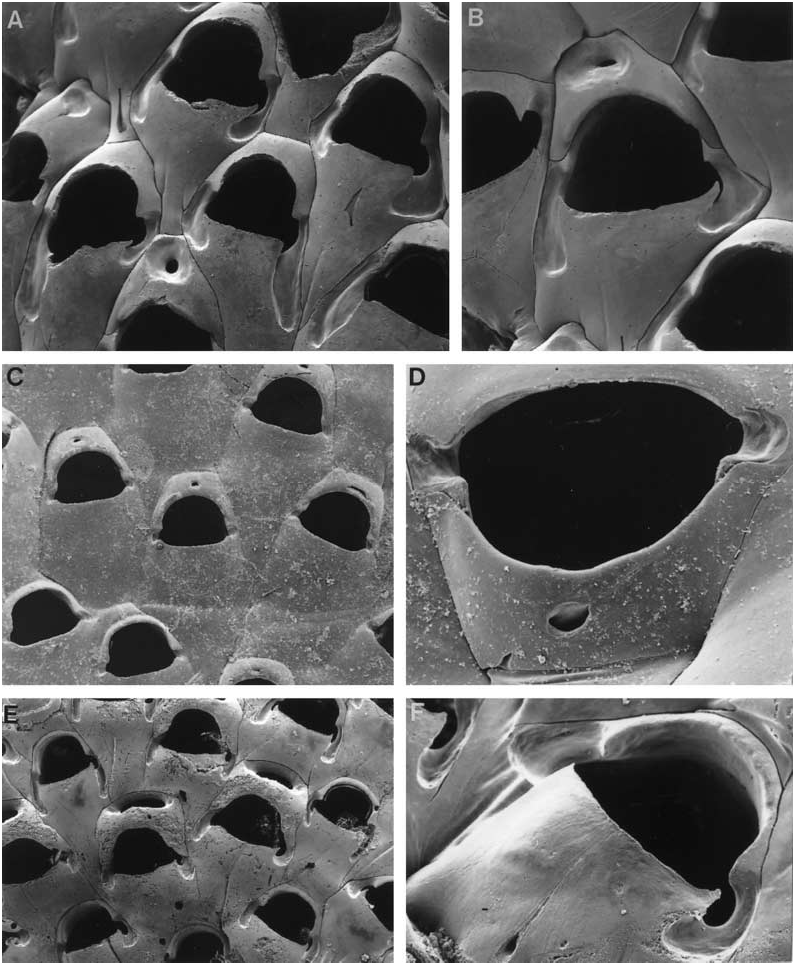
( FIG. 2A, B View Figure 2 )
Material examined
Holotype: ZIHU 02034 View Materials , from 43°3¢N, 140°35¢E, 0 m, the rocky tidal flat of Kamekawa, Kikonai-cho, Shirabeshi Prefecture , Hokkaido.
Paratype: ZIHU 02035 View Materials , same locality as holotype .
Description
Colony encrusting, multiserial, unilaminar. Autozooids contiguous, quincuncially arranged, 0.39– 0.75 mm long (0.52 ± 0.08 mm), 0.33–0.45 mm wide (0.39 ± 0.03 mm). Gymnocystal frontal shield smoothsurfaced, somewhat convex, no umbo or carina, lacking fenestrae. Orifice somewhat hat-shaped, the anter high-arched with the proximal embayments rounded, the proximal rim of the poster gently and evenly concave or straight; the lateral excavations of the gymnocyst very well developed, occurring adjacent to the embayments and extending proximally, on one or both sides, for a considerable distance, often equivalent to half the zooid length. Orifice of maternal zooids dimorphic, though not always obviously so (0.26–0.31 mm wide at the proximal margin compared to that of autozooids 3/4 0.20–0.26 mm); distal kenozooidal chamber forming a somewhat triangular cap, with a transverse to circular sloping shelf of interior wall and a small circular foramen. No basal porechambers, interzooidal communications comprising a row of uniporous septula along each lateral wall. Ancestrula not seen.
Etymology
From novellus (L.), diminutive of novus, new.
Remarks
Integripelta novella closely resembles the type species, I. bilabiata , which, however, is much larger in size. According to Soule et al. (1995), autozooids of I. bilabiata are 0.60–0.65 mm long and 0.50–0.55 mm wide with an orifice width of 0.30–0.32 mm (hence the widths of zooids and orifices do not even overlap in the two species). Additionally, the shallow gymnocystal excavations in I. novella are proportionately very much longer whereas the kenozooidal foramen is tiny and surrounded by a broader area of interior wall.
Apart from those sources already mentioned, Integripelta bilabiata has been recorded or described by Hincks (1882, 1884), Robertson (1908), O’Donoghue & O’Donoghue (1925, 1926), Osburn (1952), and Banta (1973). Soule et al. (1995) gave its range as Nootka Island, Alaska, through British Columbia to southern California and to Navidad Head, Mexico, at depths of 168–237 m, but it has also been recorded intertidally ( McBeth, 1971). It is also known from the Pleistocene of southern California ( Soule & Duff, 1957). Robertson (1908) described the operculum of I. bilabiata in some detail — it appeared to be two-layered and, together with the distal rim of the orifice, the whole structure appeared to be superficially bilabiate. Judging from observations on I. sextaria (below) the ‘bilabiate’ appearance obtains in dried material, in which the distal vestibular wall may be accentuated if slightly protruding. While the operculum proper may be thin, a descending cuticular rim around its periphery can give the appearance of thickness. The orificial structure of I. japonica (below) is not known.
Distribution
Known with certainty only from the intertidal zone at the type locality in Hokkaido .
INTEGRIPELTA JAPONICA SP. NOV.
( FIG. 2C, D View Figure 2 )
Eurystomella bilabiata :? Kataoka 1960: 396, pl. 41, fig. 3;? Hayami 1970: 330, pl. 36, fig. 3;? Hayami 1974: 222, pl. 69, fig. 2a, b;? Seo 1996: 301.
Material examined
Holotype: SMBL Type no. 399, 34°50.568¢N, 131°05.554¢E, 89–94 m, near the island of Mi-shima, north of south-west Honshu (Yamaguchi Prefecture) [Seto Marine Biological Laboratory Station 13-1, 29 September 1998].
Paratypes: SMBL Type no. 400, and NIWA P-1218, same locality as holotype .
Description
Colony encrusting, multiserial, unilaminar. Dried colonies hyaline. Autozooids contiguous, quincuncially arranged, 0.47–0.75 mm long (0.62 ± 0.07 mm), 0.32–0.57 mm wide (0.45 ± 0.07 mm). Gymnocystal frontal shield not elevated, no umbo or carina, smoothsurfaced, lacking fenestrae. Orifice somewhat more D- shaped than hat-shaped, the anter high-arched with the proximal corners bluntly condyle-like, the proximal rim of the poster straight or scarcely concave, the lateral corners curving obliquely distad towards the short indentations and adjacent excavations. Orifice of maternal zooids mostly not dimorphic, more or less identical in size and shape (0.30–0.36 mm wide at the proximal margin) to that of autozooids (0.24– 0.30 mm); distal kenozooidal chamber very small, not extending to the basal wall, the central foramen tiny, rounded or slit-like. No basal pore-chambers, interzooidal communications comprising a row of uniporous septula along each lateral wall. Ancestrula not seen.
Etymology
From japonica (L.) Japanese.
Remarks
Integripelta japonica View in CoL has almost certainly previously been confused with I. bilabiata . The two species appear similar, but published SEM micrographs ( Cook & Chimonides, 1981) allow comparison of the key taxonomic features, permitting ready discrimination of the two species. In I. bilabiata the autozooidal orifice is proportionately larger than in I. japonica View in CoL (almost half the size of the zooid) with larger proximolateral excavations on the edge of the gymnocyst (see Cook & Chimonides, 1981; fig. 5D, arrows), the maternal orifice is dimorphic, a low umbo and/or carina is often present, and the ‘ooecial kenozooid’ is larger with a larger foramen. In his original description, Hincks (1882) noted that the zooids in I. bilabiata were ‘short’ and that the ‘ooecium’ had a ‘large foramen’ (see also Hincks, 1884). The above synonymy is suggested by the published illustrations of specimens described by Kataoka (1960) and Hayami (1970, 1974). We have not examined these specimens, but the relative proportions of the orifice in relation to the entire zooid strongly indicate that they were I. japonica View in CoL , not I. novella View in CoL , which is somewhat intermediate between I. bilabiata and I. japonica View in CoL in the zooid length:orifice length ratio.
Distribution
Southern Japan Sea off south-western Honshu, 89–94 m (Seto Marine Biological Laboratory Collection).?Also Japan Sea, western Honshu, 151 m ( Kataoka, 1960), and Early Miocene of south-west Hokkaido and central Honshu ( Hayami, 1970, 1974).
INTEGRIPELTA SHIRAYAMAI SP. NOV.
( FIG. 3A–C View Figure 3 )
Lepralia bilabiata: Okada 1929: 24 , fig. 10, pl. 2, fig. 3; Sakakura 1935: 25, fig. 7.
Eurystomella bilabiata: Mawatari 1952: 280 .
Material examined
Holotype: SMBL Type no. 401, 33°02.326¢N, 132°05.608¢E, 80–86 m, off the island of Saiki-wan, Oita Prefecture, eastern Kyushu [Seto Marine Laboratory Stn 5, 26 September 1998].
Paratypes: SMBL Type no. 402, and NIWA P-1219, same locality as holotype .
Description
Colony encrusting, unilaminar, multiserial. Self-overgrowth not seen. Colony colour unknown. Autozooids contiguous, quincuncially arranged, 0.47–0.75 mm long (0.58 ± 0.07 mm), 0.28–0.55 mm wide (0.41 ± 0.06 mm). Gymnocystal frontal shield flat, centrally smooth and imperforate. Orifice longer than wide, somewhat dumbbell-shaped, the anter high-arched and rounded with the proximal corners somewhat condyle-like; poster not wider than anter, the proximal rim gently and evenly concave. Conspicuous crescentic slits curve proximolaterally from corners of poster; below the outer edge of each slit is a narrow shelf. Traces of these slits, paired or distally continuous, occur in incompletely formed autozooids, i.e. kenozooids. No peristome, umbones, spines, or avicularia. Orifice of maternal zooids larger overall than in autozooids, the distal kenozooid with a large, transversely elongate foramen sloping distad. Interzooidal communication via tiny uniporous mural septula. Ancestrula not known.
Etymology
After Professor Yoshihisa Shirayama, director of the Seto Marine Laboratory, Shirahama, Kii Peninsula, in recognition of his contributions to biodiversity appreciation in Japan.
Remarks
This is a very striking species, easily recognizable by the crescentic lateral slits, which are very conspicuous in dead zooids. Okada (1929) attributed specimens in his collection from Mutsu Bay to Eurystomella bilabiata but illustrated the slits and, in one zooid, a spine-like umbo, described in the text as keel-like or carinate and restricted to older zooids. None of the specimens we have examined show this latter feature. Eurystomella shirayamai is also distinguished on the basis of the ‘ooecial kenozooid’, the foramen of which is larger and more bean-shaped than in E. bilabiata .
Cook & Chimonides (1981) noted the crescentic lateral slits illustrated by Okada (1929) and Sakakura (1935) and also discovered them in a specimen in The Natural History Museum, London ( BMNH 1885.8.29.1) from “Sio-u-whu Bay”. [This name, not found in modern atlases, refers to a locality south of Vladivostok, Russia, in the Japan Sea. The coordinates on the label give the following data: “Sio-u- imolateral corner of an autozooidal orifice, ¥930. whn(u) Bay, Gulf of Tartary, 42°N, 133°S ”.] Cook & Chimonides (1981) wrote: “Some of the Japanese populations have been reported to show characters [sic] states which vary somewhat from those of the eastern Pacific specimens. [The slits] are covered by brown cuticle which appears to be continuous distally with that of the operculum”. This observation accords with our interpretation that the slits are lateral extensions of the indentations seen at the proximolateral corners of most eurystomellids (see Cook & Chimonides, 1981). Unfortunately, the specimens available to us were all dead and lacked opercula and membranes. The narrow shelf below the outer edge of each slit is clearly homologous with the excavations that occur in the gymnocysts of species like I. novella sp. nov. and I. sextaria sp. nov. (cf. Figs 2A, B, F View Figure 2 and 3C View Figure 3 ). One other possibility is that the slits represent frontal foramina that have migrated laterally; this is suggested by their presence in kenozooids lacking orifices, but, in one instance, the kenozooidal slits are distally continuous and the distal part of the inverse U-shaped slit is suggestive of aborted orificial development. Further, the distalmost parts of the frontal gymnocyst merely abut, and do not fuse, with the proximal corners of the ‘ooecial kenozooid’, such that organic continuity between the operculum and the slits is possible just at or under the loci of abutment.
Distribution
Integripelta shirayamai View in CoL is endemic to east Asian waters. It has been reported (as Eurystomella bilabiata ) from numerous localities in Mutsu Bay, northern Honshu ( Okada, 1929), from south-eastern Honshu ( Mawatari, 1952), and south of Vladivostok in the Japan Sea. Pleistocene material was illustrated by Sakakura (1935), who found specimens to be very rare to common in the Dizôdô beds of the Bôsô Peninsula, eastern Honshu. Specimens examined for this paper were collected off Saiki-wan, eastern Kyushu (courtesy of the Seto Marine Laboratory).
INTEGRIPELTA UMBONATA View in CoL SP. NOV.
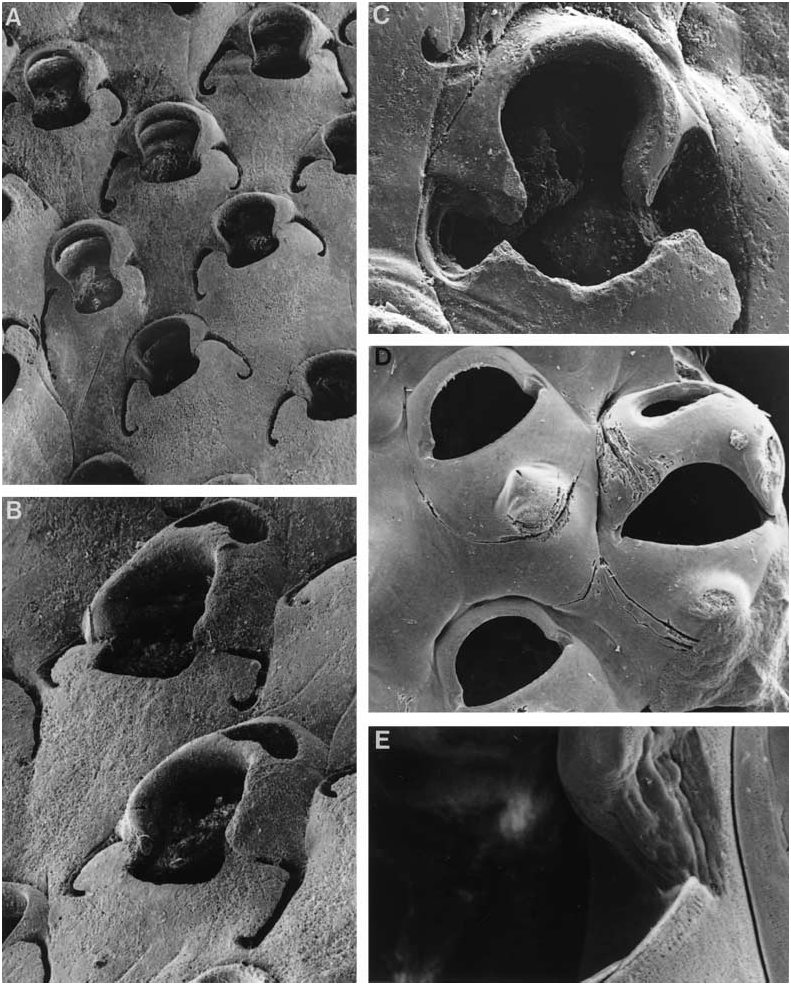
( FIG. 3D, E View Figure 3 )
Material examined
Holotype: NZOI H-751, from NZOI Stn Z 9697, 34°21.06¢S, 172°42.53¢E, 57 m, Spirits Bay , New Zealand.
Paratype: NZOI P-1219, same locality as holotype . Other material: NZOI Stns Z 9667, Z9678, Z9684, Z9687 .
Description
Colony encrusting, unilaminar. Autozooids 0.33–0.60 mm long (0.45 ± 0.006 mm), 0.22–0.49 mm wide (0.35 ± 0.008 mm). Frontal shield smooth with a conspicuous umbo in its centre. No gymnocystal foramina; no peristome, spines, or avicularia. Autozooidal orifice broad, not very high-arched, the proximal rim gently and evenly concave, not sinuous; lateral corners slightly indented and excavated. Incompletely developed autozooids are kenozooidal, with a central foramen and cuticular structure that appears to be a nonfunctional operculum. Maternal zooids obviously dimorphic, broader than autozooids with broader, somewhat D-shaped orifice (0.25–0.30 mm wide at the proximal margin; cf. 0.16–0.24 mm in autozooids). Ooecium-associated kenozooid moderately well developed frontally, with a single large foramen facing distad. Interzooidal communications via rows of tiny uniporous septula. Ancestrula unknown.
Etymology
From umbo (L.) boss, protuberance, alluding to the umbonate frontal shield.
Remarks
Integripelta umbonata is the second-known frontally imperforate New Zealand species, the other being Z. crystallina , from the Kermadec Ridge, which is biserial and its zooids lack umbones. Both species lack basolateral pore-chambers but the proximolateral rim of the orifice in Z. crystallina is somewhat elevated and has a median suture.
Distribution
Endemic to New Zealand – known only from Spirits Bay, northernmost North Island, 29–57 m depth.
INTEGRIPELTA SEXTARIA SP. NOV.
( FIG. 2E, F View Figure 2 )
Material examined
Holotype: NZOI H-750, from NZOI Stn Z 9700, 34°22.88¢S, 172°39.71¢E, 54 m, Spirits Bay , New Zealand.
Paratype: NZOI P-1220, separated part of holotype colony, same locality .
Description
Colony encrusting, multiserial, unilaminar. Autozooids contiguous, quincuncially arranged, 0.47– 0.75 mm long (0.62 ± 0.07 mm), 0.32–0.57 mm wide (0.45 ± 0.07 mm). Gymnocystal frontal shield smoothsurfaced, somewhat convex, no umbo or carina, lacking fenestrae. Orifice somewhat hat-shaped, the anter high-arched with the proximal embayments rounded, the proximal rim of the poster gently and evenly concave; the lateral excavations of the gymnocyst well developed, occurring on either side of the embayments and extending proximally a short distance. Orifice of maternal zooids obviously dimorphic (0.30–32 mm wide at the proximal margin) compared to that of autozooids (0.20–0.28 mm); distal kenozooidal chamber well developed, with a conspicuous transverse or circular foramen. No basal pore-chambers, interzooidal communications comprising a row of uniporous septula along each lateral wall. Ancestrula not seen.
Etymology
From sextarius (L.) sixth, alluding to the discovery of a sixth species of eurystomellid in New Zealand waters.
Remarks
Integripelta sextaria resembles I. bilabiata but has smaller zooids and a proportionately broader ‘ooecial kenozooid’ with a relatively very large foramen. The gymnocystal excavations are often relatively long, as in I. novella , but again, the ‘ooecial kenozooid’ and foramen are significantly larger in I. sextaria .
Distribution
Endemic to New Zealand; known only from Spirits Bay, North Island, 54 m.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Integripelta
| Gordon, Dennis P., Mawatari, Shunsuke F. & Kajihara, Hiroshi 2002 |
Eurystomella bilabiata
| Seo JE 1996: 301 |
| Hayami T 1974: 222 |
| Hayami T 1970: 330 |
| Kataoka J 1960: 396 |
Eurystomella bilabiata: Mawatari 1952: 280
| Mawatari S 1952: 280 |
Lepralia bilabiata:
| Sakakura K 1935: 25 |
| Okada Y 1929: 24 |