
Parastygocaris luisgrossoi, Peralta, Marcela, 2014
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3760.3.6 |
|
publication LSID |
lsid:zoobank.org:pub:982DAAEA-19F0-46A5-8D7F-C34E2835A60A |
|
DOI |
https://doi.org/10.5281/zenodo.6133929 |
|
persistent identifier |
https://treatment.plazi.org/id/03A287F8-BB01-9221-FF4F-2573FC883AEA |
|
treatment provided by |
Plazi |
|
scientific name |
Parastygocaris luisgrossoi |
| status |
sp. nov. |
Parastygocaris luisgrossoi n. sp.
Type Locality: stream tributary of Río de la Carpa, northern sector of Río Quinto basin, Sierra de San Luis, San Luis Province, Argentina. 32º 50’ S; 65º 59’ W, 1491 m above sea level.
Type specimens: Holotype male (FML-CRUST01109). Paratypes: five females (FML- CRUST 01110), five males (FML-CRUST01111). All collected at type locality, 6-December-2012, Col. M. Peralta, hyporheic zone.
Additional specimens examined (FML-CRUST01112): one male, four females and three immatures, same data as holotype.
Accompanying fauna: Hydra, Nematoda, Nematomorpha, Oligochaeta, Acari , Diptera ( Staphylinidae and Ceratopogonidae , larvae), Collembola, Ostracoda, Amphipoda Ingolfiellidea , Isopoda Protojaniridae, Copepoda Harpacticoida (Parastenocarididae) and Cyclopoida.
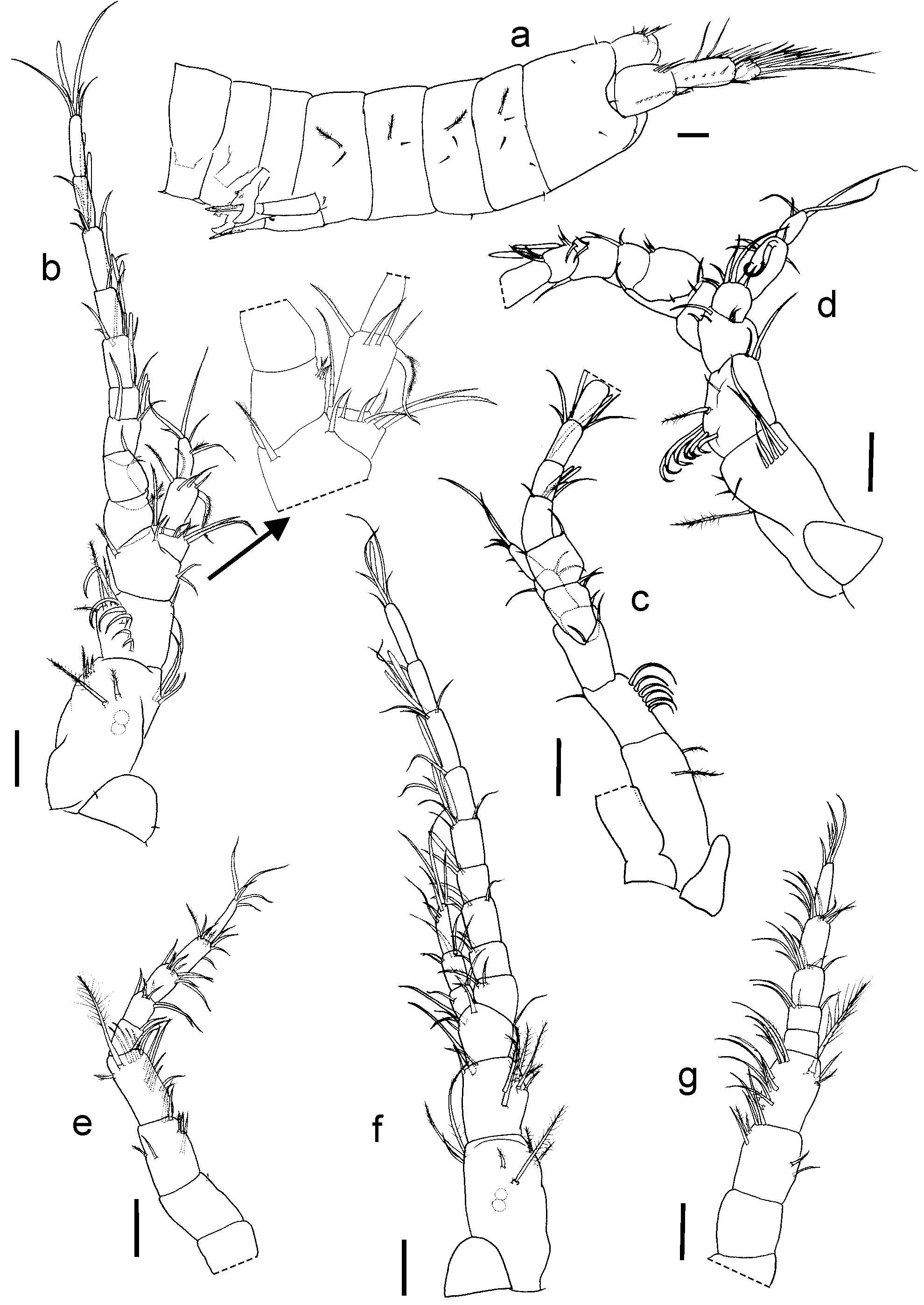
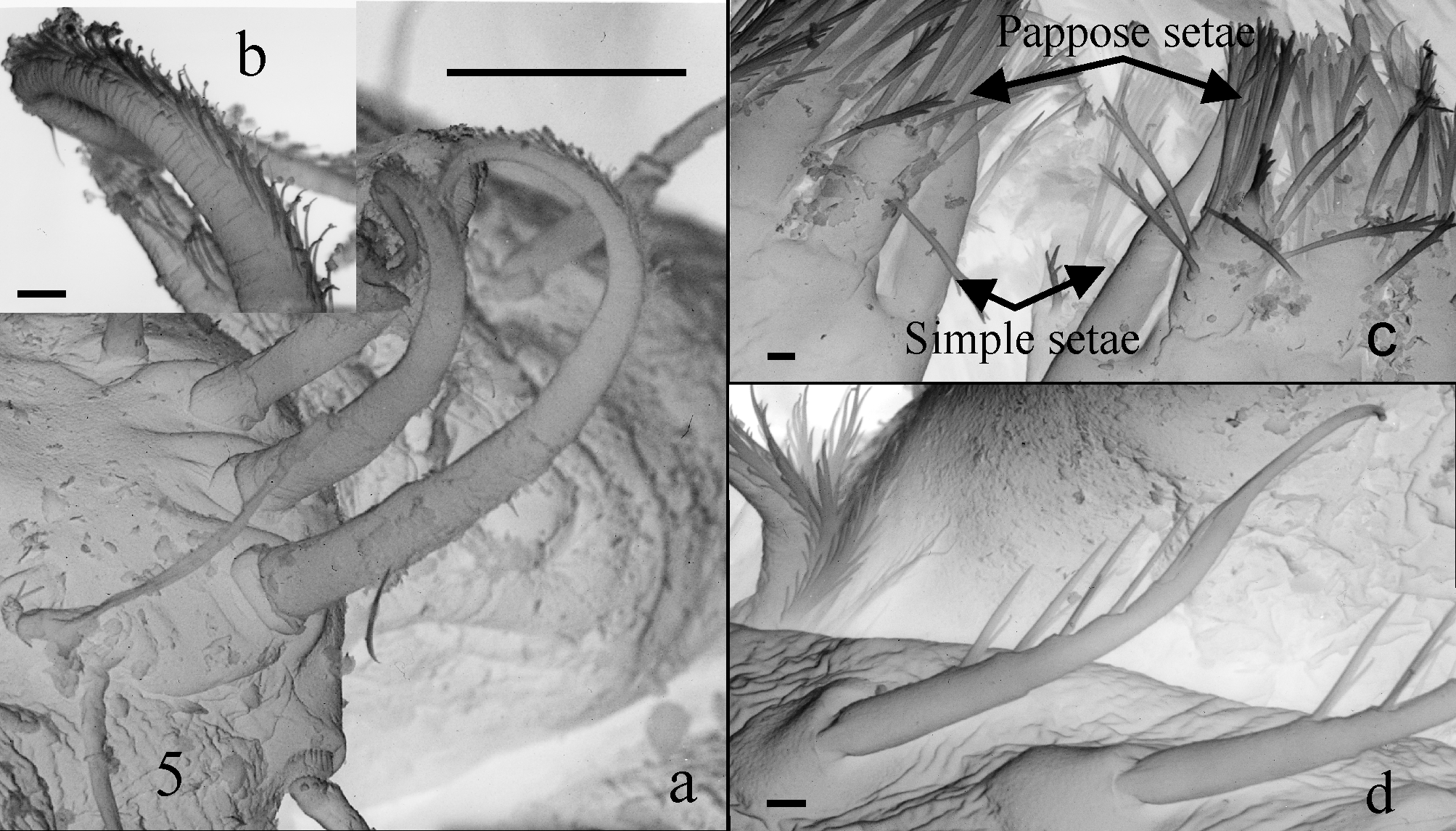
Description of Holotype. Total length 1.7 mm. Body moderately chitinized, all somites (seven pereomeres and six pleomeres) of similar size. Cephalothorax and all body somites with one or two thin simple dorsal setae at each side; second to sixth pleonites with one simple ventral seta at each side, those on second pleonite p l a c e d near to corresponding pleopod protopod; lateral margins of second to fifth pleonites ( Fig. 1 View FIGURE 1 a) each with one plumose seta (type IB1).
Rostrum ( Fig. 1 View FIGURE 1 b) trapezoidal, with blunt end and one simple seta on each lateral margin. A1 ( Figs. 1 View FIGURE 1 b, c) peduncle three-segmented, OF with nine and IF with three articles. First segment of peduncle longer than the two following segments combined, with longitudinal ridge on dorsal surface; two dorsal plumose setae (type IB1) placed near statocyst cavity; inner margin with transverse row of four–five simple setae; outer margin with lateral setiferous group composed of three subdistal plumose setae (type IB1). Second segment wider than long, outer margin with weak lateral expansion bearing three plumose setae (type IB1); dorsal surface with oblique row of six recurved ringed setae ( Fig. 5 View FIGURE 5 a) of increasing length towards distal. Third segment widened at distal end, where the two flagella are articulated; dorsal surface with one simple seta on bifurcation of both flagella; inner margin with two simple setae; distolateral angle with one plumose seta (type IA1). OF slightly curved due to morphology and articulation of proximal articles; first article wide, with inner margin slightly serrated (visible in Fig. 1 View FIGURE 1 b); bearing four short plumose setae (type IA1) (=”Sinnesorgan” Schminke 1974); second article medial margin globoid, with a deep longitudinal depression (visible in Fig.1 View FIGURE 1 d). Distal margin of third to seventh articles each with one or two aesthetascs and one–four simple setae. Ninth article with one aesthetasc and five simple setae, of which three short and two longer. IF with three articles, first article short and wide, distomedial angle with three setae (two long and one short), distolateral angle with one plumose and long seta (type IA1), dorsal surface with a transverse row of three thin short setae; second article articulating dis to- la terally to preceding article, inner margin with one recurved seta bearing fine setules on one side of shaft ( Fig. 1 View FIGURE 1 b); third article with six simple or plumose (type IA1) setae.
A2 ( Fig. 1 View FIGURE 1 e) attaining 3/5 of A1 length, with four-segmented peduncle and five-articulate flagellum; peduncle segments two to four longest, their combined length equalling flagellum length. Inner margin of third segment with three middle setae: two short plumose (type IB1), and one long simple; distomedial angle with three simple setae; outer margin with two lateral setae, one plumose (type IB1), and one simple. Surface of fourth segment with a transverse row of four distal, simple setae, and two groups of simple setae along inner margin: one, proximal, comprising two setae, other of four setae; outer margin near distal angle with two plumose setae (type IB1), one very long, and one short; distomedial angle with two simple setae. Proximal article of flagellum with one simple seta on distomedial angle; articles two to four each with six–eight distal setae, simple or plumose, those on distal margin longest. Terminal article with five simple setae on tip.
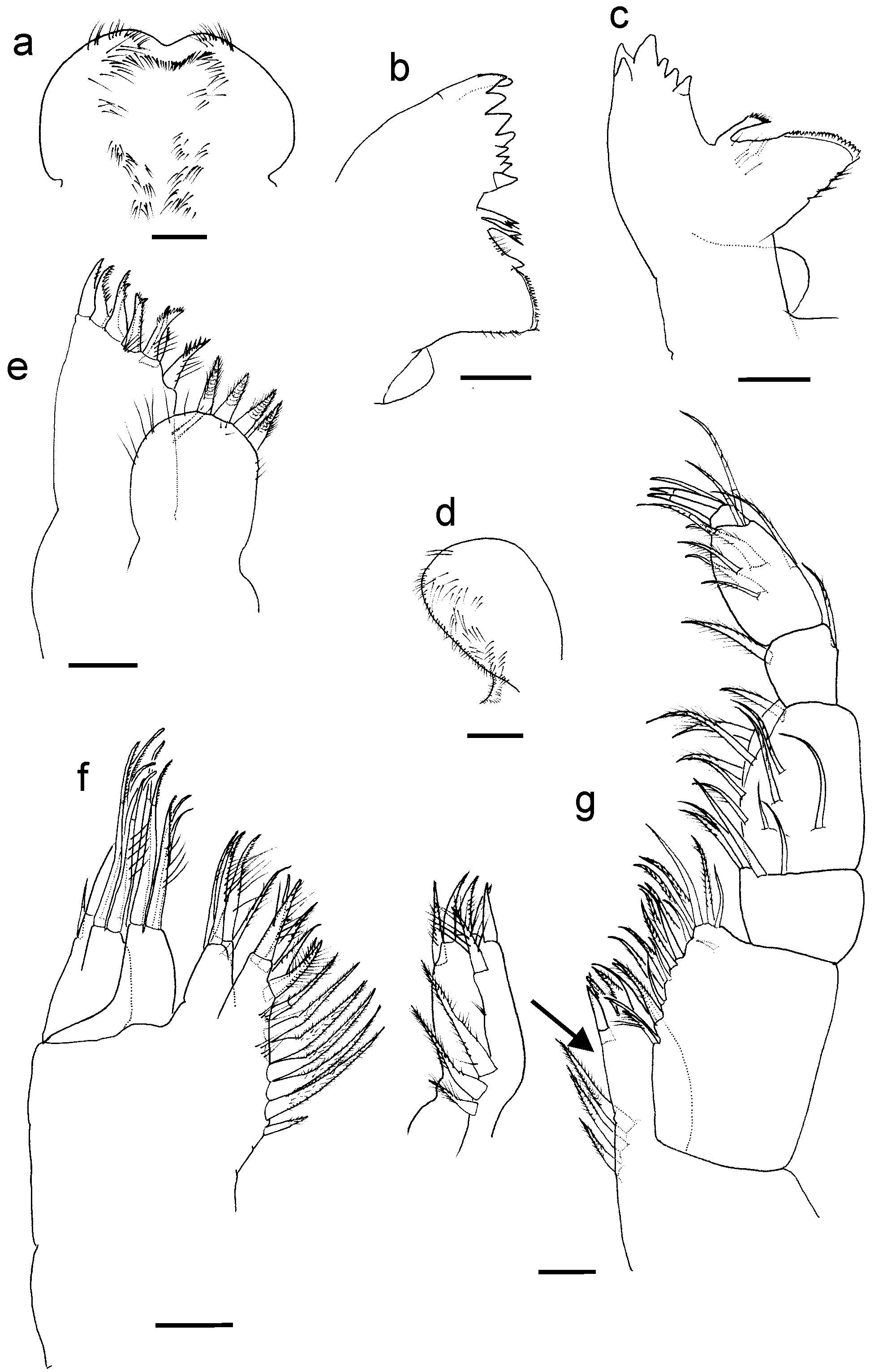
Labrum ( Fig. 2 View FIGURE 2 a): wide, with distal margin slightly excavated; anterior edge and dorsal surface with acuminate microsetae.
Left Md ( Fig. 2 View FIGURE 2 b) pars incisiva with nine conical teeth in palisade arrangement, t h e d i s t a l o n e in a plane perpendicular to remaining teeth; diastema with two serrated setae (type IIB1); margin of molar process with one spiniform process; molar process high, grinding surface surrounded by many tiny denticles. Right Md ( Fig. 2 View FIGURE 2 c) similar to left counterpart, pars incisiva with six teeth in palisade on distal curved edge: three most proximal smaller; diastema with only one serrated seta; molar process without spiniform process.
Paragnaths with oval lobes ( Fig. 2 View FIGURE 2 d); inner edge and distal margin of each lobe with long, thin acuminate microsetae.
Mx1 ( Fig. 2 View FIGURE 2 e): consisting of two endites, inner one globoid and shorter, with four strong, pappose (type IA2) setae; with lateral and ventral acuminated microsetae. Oblique end of outer endite bearing nine setae, innermost setulodenticulate (type IA4), rest arranged in double row of denticulate (type IIIA1) or setulodenticulate setae.
Mx2 ( Fig. 2 View FIGURE 2 f) with four endites (the fourth or distal endite partially subdivided), all with a doble row of distal setae; first endite bearing 13 setae: five distal plumose (type IA1), eight lateral arranged in a row, pappose (type IA2) with short setules; second endite with four distal setae (three setulodenticulate type IA4, one plumose type IA1); third and fourth endites each with five long distal setae (two simple, two setulate type IA3, one setulodenticulate type IA4); fourth endite also with one subdistal short outer seta.
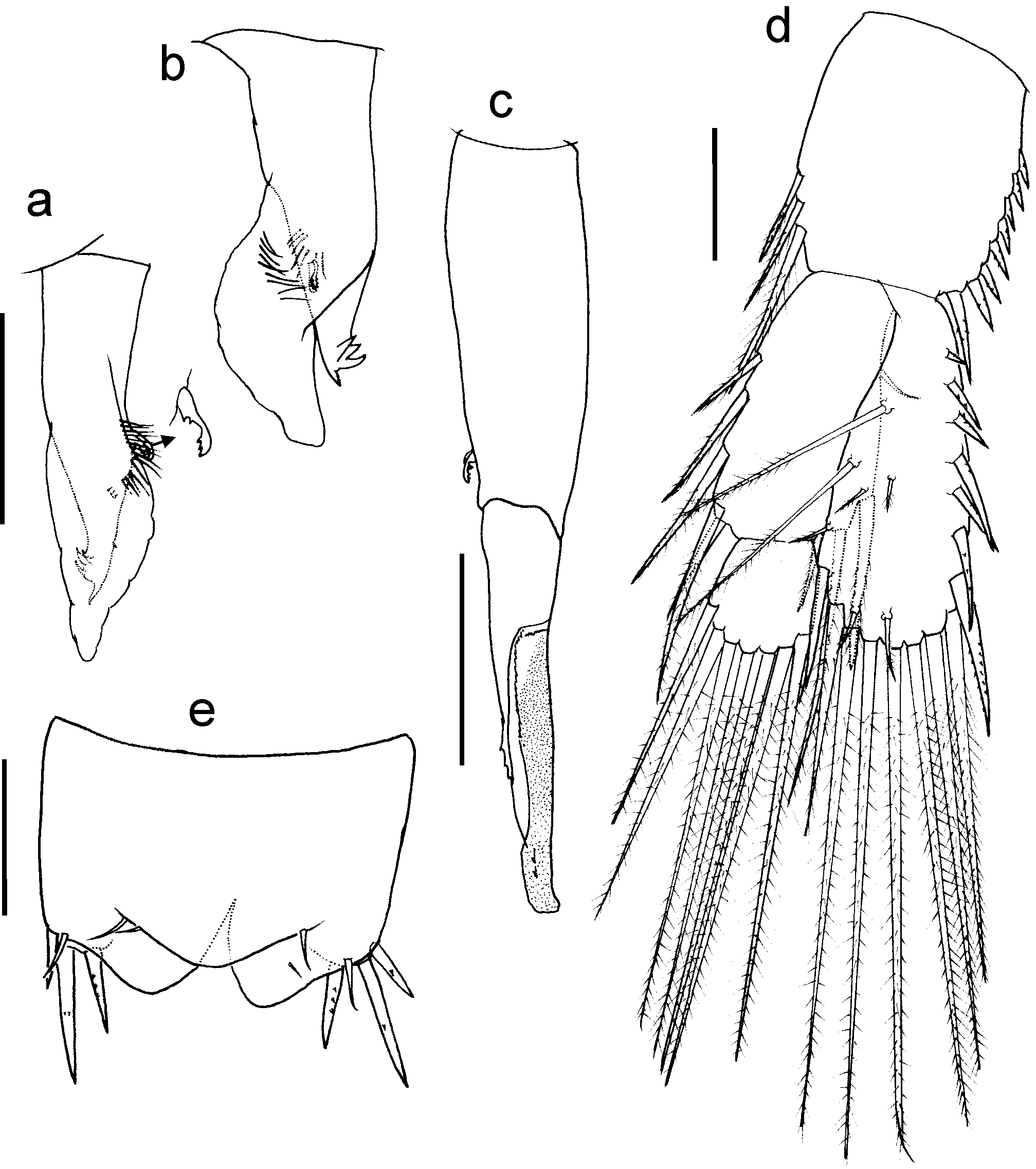
Mxp ( Fig. 2 View FIGURE 2 g) strong, seven-segmented, lacking exopod or epipodites. Coxa wide, with endite reaching half the length of basis, with nine setae: six distal ( Fig. 5 View FIGURE 5 c), of which five are pappose with setules and setuletes (type IA2), and one (innermost) is simple (type IIA1); other three pappose setae (type IA2) placed along inner margin, next to base of endite. Basis expanded, inner margin with a double row of 17 setae comprising plumose (type IA1) and simple setae intercalated. Ischium, dis tom edial angle with two plumose (type IA1) setae and two ventral, simple setae implanted close to the latter. Merus, inner margin with six plumose (type IA1) setae; ventral surface of segment with four plumose (type IA1) or simple setae. Carpus narrowest and shortest segment, both margins with one plumose seta (type IA1). Propodus, both dorsal and ventral surface with two plumose (type IA1) obliquely aligned setae, with setules and setulette on the setal shaft ( Fig. 5 View FIGURE 5 d); distomedial and distolateral angles with plumose setae (type IA1), and one seta midway of outer margin. Dactylus armed with two strong claws and one thin simple seta.
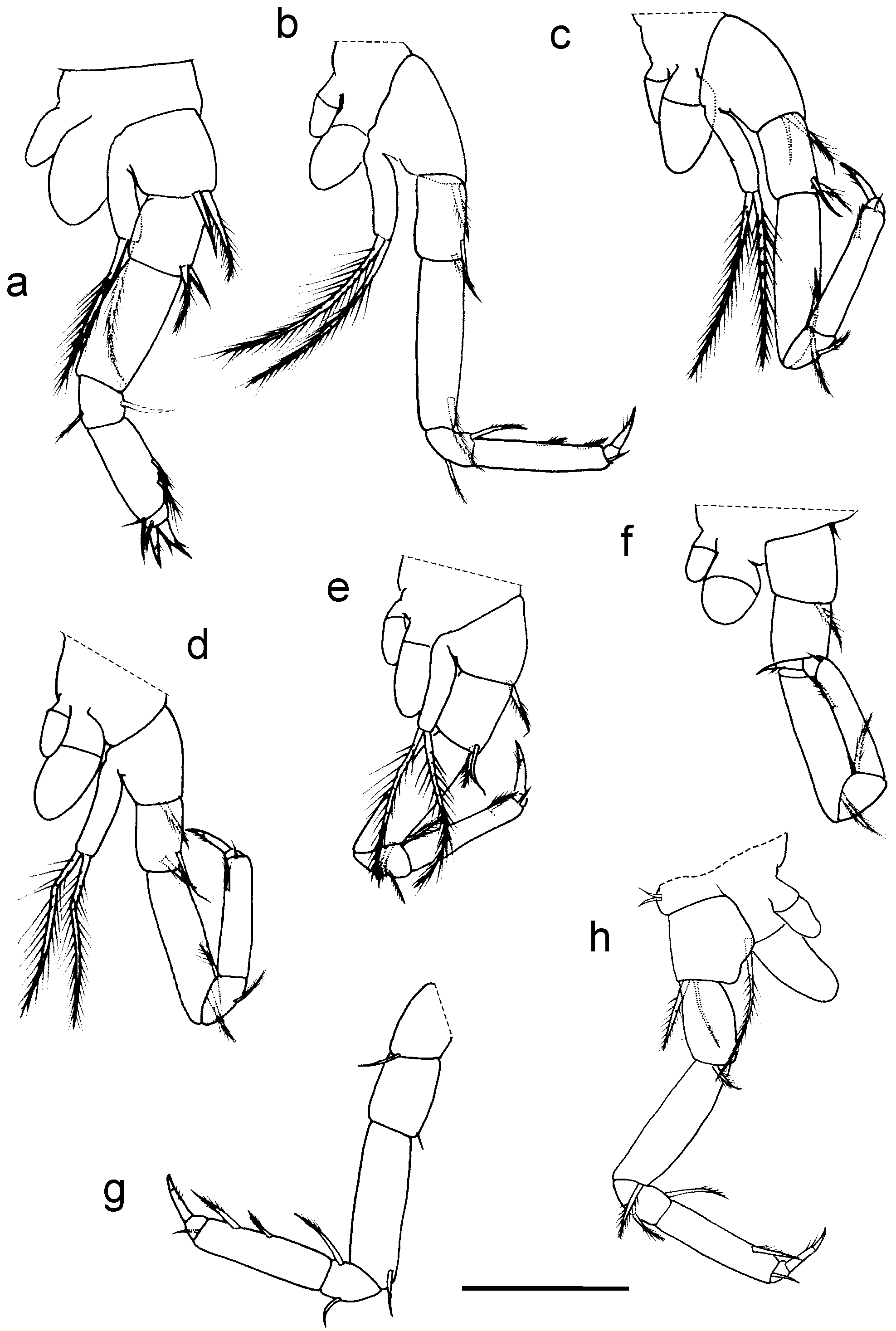
Pereopods, all seven-segmented. P1 ( Fig. 3 View FIGURE 3 a) slightly shorter and wider than rest, P2–P7 ( Figs 3 View FIGURE 3 b–g) similar in size; P1–P6: with two oval epipodites; P1–P5: basis with one-segmented cylindrical exopod bearing two long plumose setae (type IA1). P7 lacking exopod and epipodites, directed forwards. Chaetotaxy of all pereopods comprising simple or plumose setae (type IA1), basis, ischium and merus with one-two distal simple or plumose setae, carpus with one (the posterior) or two (both anterior and posterior) plumose setae, P1 and P7 propodus with two plumose aligned setae on posterior margin, P2–P6 propodus with only one seta. P1 dactylus with two strong claws, rest with a single claw. P6 coxa bearing one anterior, thin simple seta.
Pleopods 1–2 transformed into petasma, both directed forwards. Pl1 ( Figs 4 View FIGURE 4 a–b) biramous with wide protopod, rami indistinctly articulated with protopod; endopod rigid, triangular, shorter than exopod, with four spiniform processes on tip: one terminal, other three curved backwards; inner margin of protopod with two coupling setae (type IIIA2); exopod of alveolar aspect, wide, globose; middle internal area on both rami with microsetae. Pl2 ( Fig. 4 View FIGURE 4 c) uniramous, styliform, two-segmented, segments similar in length; protopod bearing two or three coupling setae on distal third of inner margin; distal segment with an external lamella encompassing two thirds of distal part, covered with tiny spiniform processes and with serrated edges.
Uropod ( Fig. 4 View FIGURE 4 d): protopod longer than wide, with aligned lateral setae on both margins, inner row consisting of eight plumose setae (type IA1); outer row with three pappose setae (type IA2), distalmost longest. Endopod and exopod equal in length. Exopod two-segmented, proximal segment slightly longer than protopod and as long as 2 ½ times length of distal one; segment prov ided with five pappose setae (type IA2) on external margin; internal margin with one proximal small simple seta and three distal pappose (type IA2), stronger setae. Distal segment with seven pappose (type IA2), very long setae. Endopod one-segmented, lined with 15 setae, of which seven on inner margin shorter; all setae plumose; dorsal surface of seg m ent with seven plumose setae (type IB1), two of which very long and strong, remaining short and smaller.
Telson ( Fig. 4 View FIGURE 4 e) 1.4 times wider than long. Anal operculum slightly convex and extending almost until tip of lateral lobes, with one or two simple small setae at each side. Lateral lobes each with three card setae (type IIB2), of which middle one strongest, plus one or two simple tiny setae.
Description of female Paratype FML-CRUST01110d. Total length: 1.69 mm (excluding antennae). A1 OF with nine and IF with three articles ( Fig. 1 View FIGURE 1 f), A2 with four-segmented peduncle and five-articulate flagellum ( Fig. 1 View FIGURE 1 g), pleopods absent. Morphology and chaetotaxy of body and appendages similar to holotype except for some characters of antennae, P1, P6 and Ur as detailed: second segment of A1 peduncle with two dorsal plumose setae (type IA1) (holotype with oblique row of six ringed setae); A1 OF straight, basal articles similar to each other (without the curvature or depression present in the A1 of holotype), third article without aesthetascs; A1 IF articles narrower and with inner setae smaller than in holotype, the second one with two thin simple setae on inner margin (without the pappose seta present in holotype); A2 fourth segment with three plumose setae near distal external angle (two in holotype), first article of flagellum without setae (one in holotype); P1 propodus with three plumose aligned setae on posterior margin (two in holotype); P6 ( Fig. 3 View FIGURE 3 h) basis with one long plumose medial seta on inner margin (absent in holotype) and two anterior simple setae on coxa; Ur protopod, inner margin with seven aligned plumose setae (eight in holotype), proximal segment of exopod with four pappose setae on external margin (five in holotype). Telson and mouthparts as in holotype.
Variability. Mean length was 1.97 mm for five male paratypes and 1.75 mm for five female paratypes. Maximum length was 2.4 mm for males and 1.90 mm for females.
Males: some variations were observed in the number of articles and chaetotaxy of outer flagellum of A1, claw of Mxp, chaetotaxy of P1 and P6: in some males (length between 1.86–2.4 mm): A1 OF with 10 articles (nine in the holotype); peduncle chaetotaxy: first article, with transverse subdistal row of five setae (four in the holotype); second article, dorsal oblique row of five ringed recurved setae (six in the holotype). In one male, the dactylus of Mxp was armed with only one claw. We observed males with three plumose aligned setae on posterior margin of P1 propodus (two in holotype) and two distal setae on P6 merus (one in holotype).
In two females (lengths 1.9 mm and 1.6 mm), the A1 IF had only two articles.
Etymology. The new species is named in honor to Dr Luis E. Grosso, Fundación Miguel Lillo-CONICET (Tucumán-ARGENTINA), one of the pioneers of biospeleology in Argentina.
Diagnosis of Parastygocaris luisgrossoi n. sp. The new species is distinguished by the following combination of characters:
Antennule outerr flagellum with nine (in females) or 9–10 (in males) articles and inner flagellum with three (in males) or two–three (females) articles. In both sexes antenna with four-segmented peduncle and fivearticulate flagellum.
Maxilla setiferous row of eight pappose setae along inner margin of internal endite. Maxilliped basis inner lobe with 17 setae arranged in a double row.
Uropod protopod, inner margin with seven–eight plumose setae; endopod with a total of 15 marginal setae, seven of which on the inner margin; exopod, distal segment lined by seven pappose setae. Petasma: pleopod 1, inner ramus triangular, stiff, tip with spiniform processes and coupling setae. Pleopod 2 styliform, with two segments of similar length; distal segment with an external lamella and serrated edges.
Secondary sexual characters:
Male: antennule, second article of peduncle with an oblique row of five–six recurved ringed setae; outer flagellum with basal part slightly curved due to morphology and articulation of its articles, second article globular, with internal depression; second article of the inner flagellum articulating disto-laterally to preceding article.
Female: pereopod 6 with one long and strong plumose seta on basis.
Systematic comments. The Stygocarididae are the most differentiated anaspidacean family, and the sister group of Patagonaspididae ( Schminke 1975; Schram 1984; Grosso & Peralta 2002). A morphological simplification of appendages (short antennae; Md, Mx1, Mx2 each devoid of palp; Mxp without epipodites or exopod; exopods of pereopods one-segmented or absent; pleopods vestigial or absent) is evident in both families, and especially marked in the species of Stygocarididae .
The Stygocarididae View in CoL includes four genera: Parastygocaris Noodt, 1963 , Stygocaris Noodt, 1963 View in CoL , Stygocarella Schminke, 1980 View in CoL , and Oncostygocaris Schminke, 1980 . An analysis of the diagnosis of Parastygocaris ( Noodt 1970) shows that the one-segmented cylindrical exopod on P1–5 represents its only generic distinctive feature, the rest of characters mentioned as dia g nos tic being shared with some Stygocaris View in CoL species. The only known species of Patagonaspididae , Patagonaspides sandroruffoi Grosso & Peralta, 2002 shares with Parastygocaris the presence of one-segmented exopods on P1–5; however, both genera differ in the structure of the exopod: it is cylindrical with 2 setae in Parastygocaris whereas truncated and ribbon-shaped, with 8 setae in Patagonaspides Grosso & Peralta, 2002 .
When comparing Parastygocaris luisgrossoi n. sp. with related taxa species, we considered the following characters: rostral morphology; articles and secondary sexual characters of antennae; features of the pleopods; and chaetotaxy of Mx2, Mxp, Pl, and Ur (Table 1).
The new species is similar to P. g o e rs s i by several characters: number of articles of A1 OF; A1 IF of male with secondary sexual characters; number of inner setae on the first endite of Mx2; number of marginal setae on Ur protopod; and morphology of the rostrum.
P. luisgrossoi n. sp. shares with P. c l a p s i the number of setae on the Mxp endite and the chaetotaxy of the Pl1inner ramus.
Regarding pleopod morphology, all Parastygocaris species are greatly similar except for P. clapsi due to its trisegmented Pl2.
Within the genus Parastygocaris , the new species is small (1.97 mm mean length for males; 1.75 mm for females) taking into account that P. andina , the largest Stygocarididae View in CoL species, exceeds 4 mm long. Besides being the largest species, P. andina has several unique characters: it has the longest A1 and the greatest number of cuticular extensions in many appendages (Mx2, pereopods, Pl2, Ur).
Comments on setal types. The study of the chaetotaxy of Parastygocaris luisgrossoi n. sp. under optical and electron microscopy has allowed the distinction of a new type of ringed seta as well as 15 types of previously described cuticular extensions ( Peralta 2010):
- Macrosetae Type I: with infracuticular articulation (IA1: plumose; IA2: pappose; IA3: setulate; IA4: setulodenticulate a and setulodenticulate b); and with supracuticular articulation (IB1: plumose)
- Macrosetae Type II: shaft smooth (IIA1: simple; IIA3: aesthetasc); and with denticles (IIB1: serrate; IIB2: card)
- Macrosetae Type III: with denticles (IIIA1: denticulate; IIIA2: coupling seta); and without denticles, strong (IIIB)
- Microseta Type A: acuminate
- Spiniform processes
The new species shows a remarkable enhancement of the sensory function of the male A1, where a new seta type, here called ringed, occurs and which had not been detected before in any Stygocarididae species. This element is a macroseta with infracuticular articulation (type I), circular in cross-section, characterized by parallel tenuous rings around the shaft and one fringe of multiple fine short setules arising from the shaft wall ( Fig 5 View FIGURE 5 a). An oblique row of five–six of these recurved setae occurs only on the dorsal surface of the second segment of the peduncle of A1. These setae resemble those inserted on the A1 IF of Anaspides tasmaniae Thomson , 1 8 9 3, an exclusively male character. However, in A. tasmaniae , these setae do not bear setules on the shaft, but small scale-like plates.
In addition, the A1 IF of the male of Parastygocaris luisgrossoi n. sp. has a recurved seta with fine setules on one side of its shaft; thus, this setal type is a special type of pappose seta (type IA2).
In the Mxp, an appendage subjected to high mechanical stress, the cluster of robust distal setae of the coxal endite includes only one simple seta (type IIIB) ( Fig. 5 View FIGURE 5 c), whereas the rest are pappose. This simple seta is placed on the inner angle of the endite. Further detailed studies of other Stygocarididae species are necessary to ascertain whether this character is an autapomorphy.
Biogeographic remarks. The evolution of the Anaspidacea is believed to have occurred only on their Gondwanan refugium ( Schram & Hessler 1984). Stygocaris (Stygocarididae) has a disjunct geographical distribution in Australia, New Zealand, and South America (Chile- Argentina). This link across the southern Pacific is a manifestation of an ancient dispersal pathway, or alternatively a vicariant track, that included Antarctica when Gondwana was intact ( Schram 2008), and thus, the generic differentiation w i thi n the fa m ily must have occurred at least before the Cretaceous ( Grosso & Peralta 2002), when the breakup of Gondwana accelerated, and before the uplift of the South American Andes.
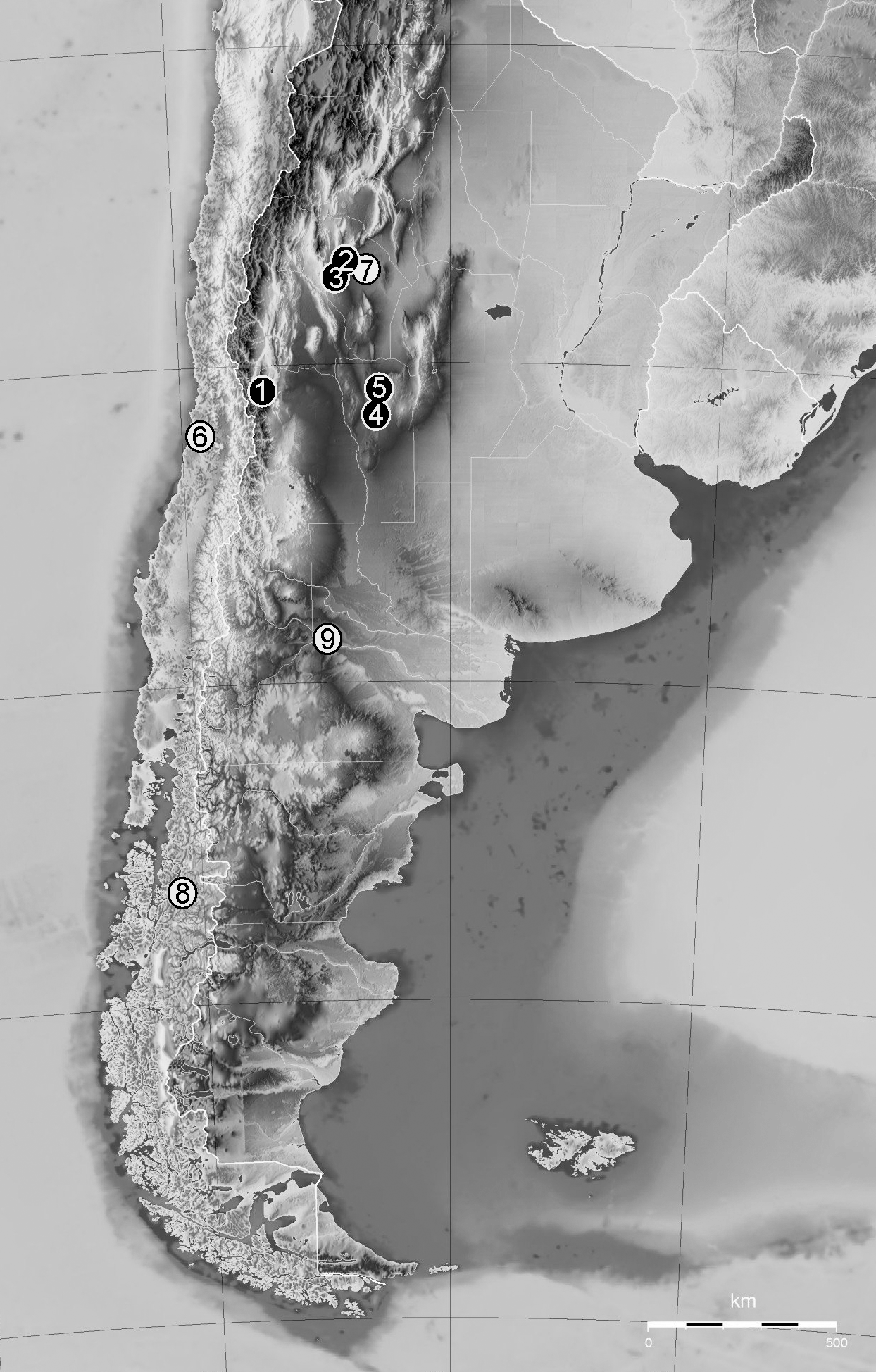
The new Stygocarididae species, was collected at the southern Sierra de San Luis ( Fig. 6 View FIGURE 6 ), a unit that is part of the eastern Sierras Pampeanas characterized by a base of Precambrian to Paleozoic rocks, partially covered by younger sequences ( Brodtkorb et al. 2009). Some geologists consider that the Sierras Pampeanas represent one of the fragmented remnants of Gondwanan paleosurfaces, geomorphological systems that were generated when the former Gondwana super-continent was still in place and whose preservation was possible due to similar tectonic conditions in the drifting fragments ( Rabassa et al. 2010).
The rise of the Andes was a pivotal event for the ancient freshwater biota of the region, because it created the current Magdalena and Paraguay Rivers and most of the Orinoco System, produced changes in former river courses ( Lundberg et al. 1998, Potter 1997), and also created new upland habitats (eg. Sierras Pampeanas intermountain valleys). All these events must have influenced the modern pattern of distribution of the stygobitic Stygocarididae in South America; in addition, the region was affected by repeated marine incursions from the Pacific, South Atlantic and Caribbean since Paleozoic times. All these happenings are potential vicariant events for groundwater biotas.Taking into account the above mentioned factors, as well as the long evolutionary history of Anaspidacea , we postulate the probable existence of an ancient groundwater basin in what is now the intermountainous valleys of western South America south of 28ºS, which would have harboured interstitial fauna of great and diverse antiquity. The ocurrence of Ingolfiellidae amphipods, a group that presumably dates from at least Triassic times ( Vonk & Schram 2003), and of Gondwanan Protojaniridae isopods as accompanying fauna of the new Parastygocaris species, supports this hypothesis.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Parastygocaris luisgrossoi
| Peralta, Marcela 2014 |
Patagonaspides sandroruffoi
| Grosso & Peralta 2002 |
Patagonaspides
| Grosso & Peralta 2002 |
Stygocarella
| Schminke 1980 |
Oncostygocaris
| Schminke 1980 |
Parastygocaris (
| Noodt 1970 |
Parastygocaris
| Noodt 1963 |
Stygocaris
| Noodt 1963 |