
Rhadinella dysmica, Campillo, Gustavo, Dávila-Galavíz, Luis Fernando, Flores-Villela, Oscar & Campbell, Jonathan A., 2016
|
publication ID |
https://doi.org/10.11646/zootaxa.4103.2.6 |
|
publication LSID |
lsid:zoobank.org:pub:6FE194B5-091E-4CBE-A105-7A72A7D445E5 |
|
DOI |
https://doi.org/10.5281/zenodo.5682458 |
|
persistent identifier |
https://treatment.plazi.org/id/03A387A4-3F4E-FFBB-2FB5-FAB2FBD97117 |
|
treatment provided by |
Plazi |
|
scientific name |
Rhadinella dysmica |
| status |
sp. nov. |
Rhadinella dysmica View in CoL sp. nov.
Figs. 1–2 View FIGURE 1 View FIGURE 2 , Table 1 View TABLE 1
Holotype. Adult female, ENCB 18951, obtained 17 July 2014 by L. F. Dávila Galavíz near cueva de Tepozonales ( 17.2853 N, – 99.3662 W; 432 m above sea level, asl hereafter), Cuajilotla, 20 km S Mochitlán, Guerrero, Mexico. The individual was found near the entrance of the cave, crawling on the ground.
Diagnosis. A member of Rhadinella , genus characterized by gracile body, head only slightly distinct from neck; inverted dark U-shaped marking on rostral; anterior supralabials usually with dark edges and pale centers; conspicuous or at least evidence of pale collar on neck; with reduced to small supracloacal tubercles in adult females; no subpreocular scale; posterior (2–7) teeth larger and thicker than preceding teeth, all enlarged teeth on same plane, no broad diastema; with basal nude pocket on hemipenis. Rhadinella dysmica is further characterized by the combination of a distinctive white collar, undivided at middorsum, extending 2.0–2.5 scale lengths at middorsum and 3–4 scale lengths on side of head; temporals 1 + 2; smooth dorsal scale rows 17, unreduced throughout body; ventrals 172; cloacal scute divided; divided subcaudals incomplete 63+N; dorsal coloration dark brown to blackish, almost totally obfuscating longitudinal striping, ground color barely extending to lateral edges of ventrals.
Rhadinella dysmica may be distinguished from many congeners (i.e., R. godmani , R. hannsteini , R. hempsteadae , R. kanalchuchan , R. kinkelini , R. lachrymans , R. montecristoi , R. pegosalyta , R. rogerromani , R. serperaster , and R. tolpanorum ) by dark dorsal body coloration and absence of conspicuous striping. Additionally, the new species differs from R. godmani , R. hempsteadae , R. kanalchuchan , R. montecristoi , R. pegosalyta , and R. serperaster by having 17 dorsal scale rows (vs. more than 17 dorsal scale rows).
Rhadinella dysmica is morphologically similar to other species having mostly uniformly blackish dorsum, with or without indistinct dark vertebral line and pale dashes on some dorsal scales (i.e., R. schistosa , R. donaji , R. pilonaorum , and R. posadasi ). In R. dysmica the color and configuration of the collar alone diagnostic from all other species of dark Rhadinella , extending from about posterior tips of parietals for several scale lengths, being white, and distinctly set off from mostly black head-cap. In R. shistosa collar U-shaped and usually broken medially ( Fig. 3 View FIGURE 3 ); in R. donaji white coloration of collar extends on to head, involving most of parietals and part of frontal; in R. pilonaorum collar orange with top of head mostly auburn mottled with black; in R. posadasi the collar is dark orange or auburn. Other features distinguishing these dark-colored species of Rhadinella include fewer ventrals ( 151–156 in females), fewer subcaudals ( 31–34 in females), and 1+1 temporals in Rhadinella shistosa ; more subcaudals ( 100–111 in females) in R. pilonaorum ; and fewer ventrals ( 143–146 in females) and more subcaudals ( 86–93 in females) in R. posadasi . Fourth and fifth infralabials contact posterior pair of genials in R. schistosa , R. pilonaorum , and R. posadasi (vs. the third and fourth in R. dysmica and R. donaji ); distal portion of tails incomplete in holotypes of R. dysmica (a female) and R. donaji (a male), however with 66 subcaudals R. dysmica appears to have higher number than R. donaji , which has 40.
Description of the holotype. Adult female with few small supracloacal tubercles; head length 9.5; head width 4.0; neck 3.0; snout-vent length 256; tail missing perhaps no more than 2–3 distal subcaudals, length 85 + N; eye diameter 1.1; naris to anterior edge of eye 3.0; head scales arranged in typical colubrid fashion with paired internasals, paired prefrontals, broad median frontal, narrow paired supraoculars, and elongate paired parietals; internasal suture 0.6, about 33% length of prefrontal suture; frontal length 3.0, about 120% as long as its distance from tip of snout, 67% as long as parietals; interocular distance 3.5, 63% occupied by frontal; mental broad, 1.5 x 1.0; two pairs of genials, subequal in size, posterior pair wing-shaped, deeply incised along median suture. Dorsal scales smooth, in 17/17/17 rows; four rows at midlength of tail; apical pits appear to be absent; supralabials 7/7, third and fourth entering orbit; infralabials 7/8, first pair in contact behind mental along midventer, first to third/ first to fourth in contact with anterior genials, respectively; third and fourth/fourth and fifth infralabials in contact with posterior genials, respectively; preoculars 1/1, subpreocular absent; postoculars 1/1; temporals 1+2; preventrals 2; ventrals 172; cloacal plate divided; subcaudals 63+N (incomplete), divided; pupil round.
Left maxilla bearing 12 teeth, increasing in size posteriorly to ninth tooth, with 4–5 teeth in middle of series noticeably enlarged, several posteriormost teeth smaller; tooth nine even with anterior edge of ectopterygoid process; teeth arranged in straight line except for slight medial curvature of anterior of maxilla; no tooth noticeably offset; no pronounced diastema within series.
In life, black head cap extending from internasals to near posterior edges of parietals, including prefrontals, frontal, supraoculars; rostral with inverted, black U-shaped marking bordering lingual groove, bordered above with dusky white on edges of first supralabials, anterior nasals, and internasals; very small amount of beige mottling on posterior nasal, loreal and anterolateral portion of prefrontal; parietals with pale spot lying to either side of common parietal suture; lateral and posterior edges of parietals with dusky white borders; side of head including nasals, loreal, preocular, and postocular blackish; upper edges of most supralabials blackish, whitish below, with lower white area of supralabials sometimes with dark brown to blackish spotting; most of primary, secondary, and upper tertiary temporals with distinctive dark brown borders and whitish centers; mental with dark brown lateral borders; infralabials 1–5 with irregular dark brown vertical streaks either centrally or along suture; anterior one-third to onehalf of anterior genial with dark brown pigment; nuchal white collar extending from posterior edge of parietals posteriorly 2.0–2.5 scale lengths, about 3–4 scale lengths wide on side of head, involving posterior portion of ultimate supralabials; dorsal pigment dark brown, barely encroaching on lateral portion of ventrals, more so on subcaudals; first two rows of dorsals with whitish central dashes; dorsal scale rows 4–5 with inconspicuous brown stripe, bordered below by a darker brown line on adjacent portions of scale rows 3–4; scale row 6 with inconspicuous dark brown stripe extending to adjacent portions of scale rows 5 and 7; subcaudals white with irregular brown pigment along midventral area; distal one-third of tail mostly brown; iris dark bronze. In preservative (ethanol after formalin) dark pigment of head and body mostly dark brown; striping of body obscured; collar and ventrum white.
Etymology. The specific epithet is derived from the Greek ἐφέσπερος ( dysmikos), meaning western, in reference to the westernmost distribution of this species of the genus. We propose standard English and Spanish names of Guerrero Slender Leaf Litter Snake and Hojarasquera Delgada de Guerrero, respectively.
Distribution and habitat. The type-locality lies at 432 m asl at the edge of a small valley occupied the small village of Cuajilotla ( Fig. 4 View FIGURE 4 ) in the foothills of the eastern portion of Sierra Madre del Sur in Guerrero in tropical deciduous forest. The predominant types of vegetation at higher elevations in the region are oak forest and pine-oak forest, grading into tropical deciduous forest, which is restricted to the lowlands of foothills, valleys, and slopes ( Leopold 1950). The type-locality ( Fig. 5 View FIGURE 5 ) is drained by the Río Chapolapa, an upper tributary of the Río Omitlán- Azul, which flows into the Río Papagayo.
Other species of amphibian and reptiles recorded between 340–432 m in the vicinity of the type-locality were Rhinella marina, Craugastor rugulosus, Agalychnis dacnicolor, Smilisca baudinii, Anolis dunni, Aspidoscelis guttata, Holcosus undulatus, Phyllodactylus lanei, Phyllodactylus tuberculosus, Ctenosaura pectinata, Sceloporus siniferus, Urosaurus bicarinatus, Boa imperator (sensu Hynková et al., 2009), and Leptodeira annulata .
The taxonomic histories of snakes placed in the genus Rhadinella were most recently discussed by Myers (2011) and Campbell (2015). Most of the species in the genus occur in Nuclear Central America, with a number of them inhabiting Chiapas, Mexico. Of the species having a dark, blackish dorsum, with or without inconspicuous stripes, only two species where known from Mexico until recently: R. schistosa ( Smith 1941; the generotype) from the eastern versant of Veracruz and R. posadasi from extreme southern Chiapas. The genus was seemingly absent from the Pacific versant of southern Mexico until the recent description of R. donaji Campbell (2015) from the Pacific highlands of Oaxaca. The subsequent discovery of R. dysmica at a relatively low elevation ( 432 m) in Guerrero is somewhat of a surprise. Snakes occurring at low elevations in tropical deciduous forest usually have rather extensive distributions in this widespread and continuous lowland habitat, not being affected by the isolating effects of elevation.
Many species of Rhadinella have distinctive longitudinal body striping that serves as a useful identifier (see Myers 1974: fig. 28). The pattern in R. dysmica with the combination of dark striping on adjacent portions of scale rows 3–4 and on the sixth scale row and adjacent portions of fifth and seventh scale rows is not found in any other species of Rhadinella . Unfortunately, the dorsal patterning in the dark-colored species is often difficult to discern or obscured altogether. We are able to address the striping in R. dysmica only because of the excellent photographs taken in life. Under certain light conditions, striping pattern in life is obscured owing to the highly iridescent scales of this snake, and in preservative there is a tendency for striping to become even more obscure.
The geographically closest congeners to R. dysmica are R. schistosa and R. donaji ( Fig. 5 View FIGURE 5 ). Rhadinella schistosa occurs about 300 km to the northeast, separated by the xeric Tehuacán-Cuicatlán Valley. Most individuals have been taken from near 1000 m asl in mesic tropical evergreen forest or cloud forest. Rhadinella donaji occurs about 275 km to the East and, similarly to R. dysmica , inhabits the Pacific versant, but their habitats are dramatically different: R. dysmica occurs in tropical deciduous forest at 432 m asl, whereas R. donaji is only known from pine-oak forest at 2195 m asl. The elevational distribution of these species remains to be ascertained but it is unlikely that their ranges overlap given habitat and elevational differences between the two species, and the fact that distribution of other species of Rhadinella are geographically restricted. There are several traits apparent in members of the genus Rhadinella with a dark dorsum. First, each species seems to be restricted to a particular habitat, whether it be cloud forest, pine-oak, rainforest, or in the case of R. dysmica , tropical deciduous forest. Secondly, the pale head pattern shows little variation within species ( Myers 1974). Both of these traits provide evidence for the distinction of R. dysmica from R. donaji .
The segmental counts for R. donaji and R. dysmica are high compared to other members of the genus and given their distributions it is likely they are closely related. With a sample of one for each of these species, it is not possible to address variation. Nonetheless, these species sufficiently differ from each other that their heterospecificity can hardly be doubted. Beside features outlined in the diagnosis, careful examination of these species reveals the following differences in color pattern. The most distinctive element of pattern is the bright white collar which in R. dysmica begins behind the parietals, although it is contiguous with parts of the parietals that are edged with dingy white. In R. donaji the white collar is continuous with the mostly pale parietals. In R. dysmica the temporals have mostly dull white centers with irregularly black edging and the loreal region is dark with small tan flecks, whereas in R. donaji the temporals are mostly white with irregular dark mottling and the loreal region is mostly white. The dark dorsal ground coloration barely reaches the lateral portions of the ventrals but strongly encroaches on lateral portions of the subcaudals in R. dysmica , whereas in R. donaji the dorsal coloration fails to reach the ventrals and barely encroaches on the subcaudals. In R. dysmica dark pigment along the midventer of the tail forms a grayish band, but in R. donaji the subcaudals are immaculate. The lower two scale rows in R. dysmica have very faint and irregular indications of pale dashes in their center and in R. donaji the lower fourth scale rows have distinct pale dashes which are especially conspicuous on the lower two rows.


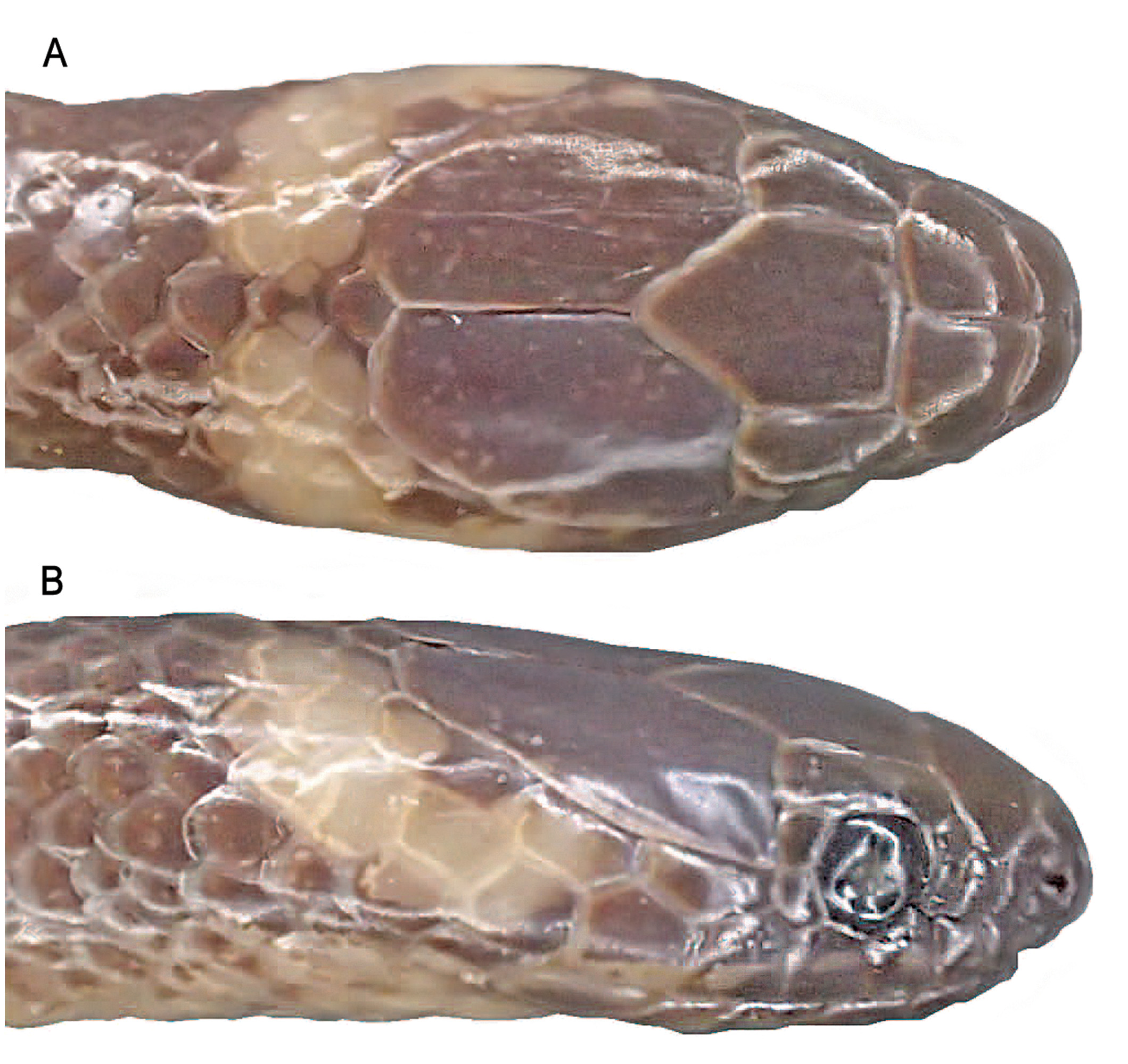


TABLE 1. Comparison of selected features from Mexican species of Rhadinella with dark-colored dorsum.
| Rhadinella dysmica | Rhadinella donaji | Rhadinella schistosa | Rhadinella posadasi | |
|---|---|---|---|---|
| Head pattern | Head mostly black followed by white collar; lateral edges of parietals dingy white | Head mostly white with scant dark mottling followed by white collar | Top of head uniformly dark followed by U- shaped white collar, usually broken medially | Top of head with uniformly dark head cap followed by orange or auburn collar |
| Infralabials contacting posterior genials | Third and fourth/ fourth and fifth | Third and fourth | Fourth and fifth | Fourth and fifth |
| Supralabials | 7 | 7 | 8 | 7 |
| Temporals | 1 + 2 | 1 + 2 | 1 + 1 | 1 + 2 |
| Ventrals | Males: unknown Female: 172 | Male: 166 Females: unknown | Males: 145–147 Females: 151–156 | Males: 136–141 Females: 143–146 |
| Subcaudals | Males: unknown Female: 63+ N | Male: unknown Females: unknown | Males: 40–42 Females: 31–34 | Male: 92–95 Females: 86–93 |
| No. maxillary teeth (sample size) | 12 (1) | 11 (1) | 16–17 (5) | 11 (2) |
| Discussion |
| ENCB |
Universidad de Autonoma de Baja California |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |