
Allodorylaimus, Andrassy, 1986
|
publication ID |
https://doi.org/ 10.1080/00222933.2023.2195565 |
|
DOI |
https://doi.org/10.5281/zenodo.7859774 |
|
persistent identifier |
https://treatment.plazi.org/id/03A387F5-FFEB-1D34-FF3F-1F37FBB2FB76 |
|
treatment provided by |
Plazi |
|
scientific name |
Allodorylaimus |
| status |
|
Taxonomy of Allodorylaimus
Diagnosis
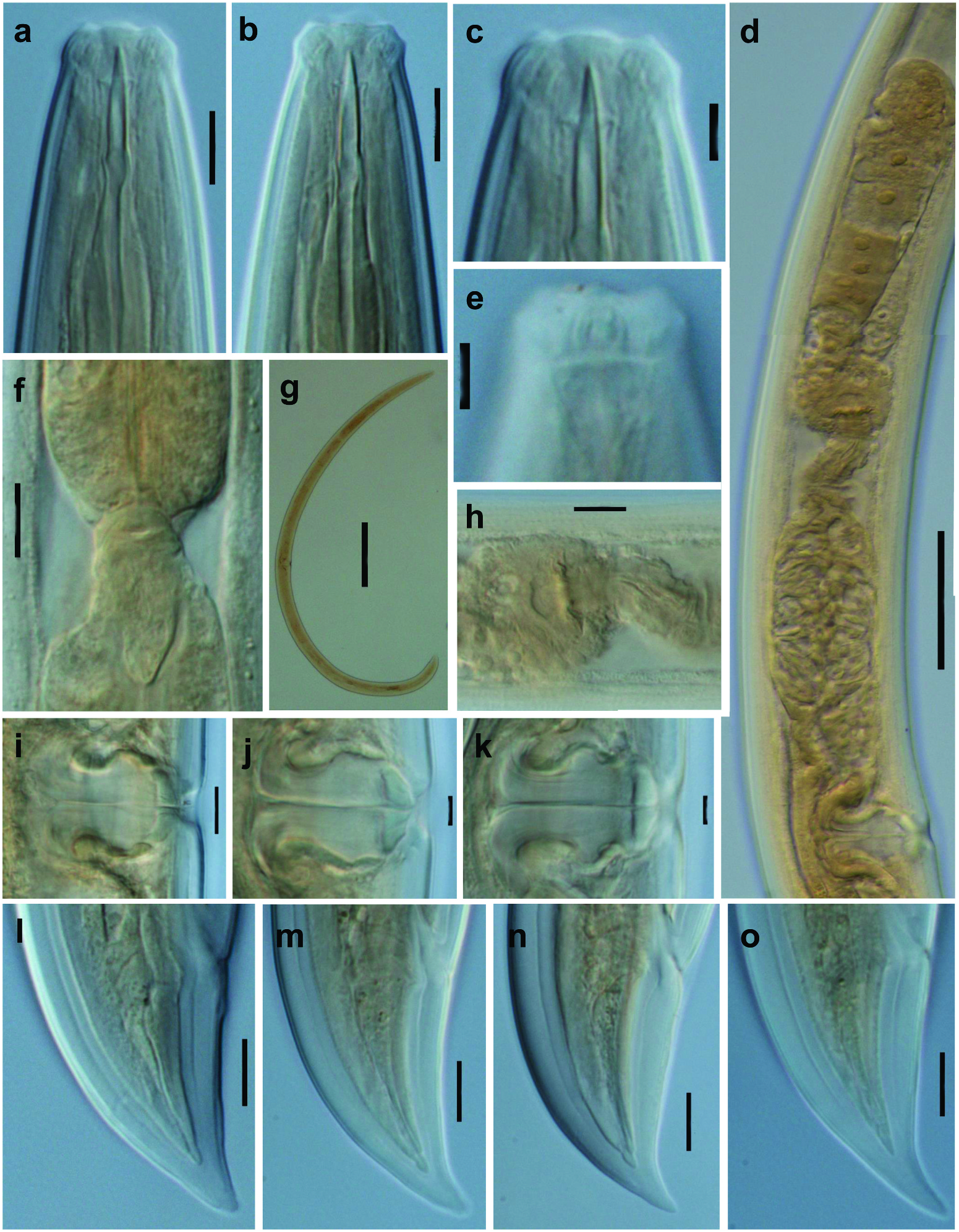
Qudsianematidae , Qudsianematinae. Small- to large-sized nematodes, 0.80–3.28 mm long. Cuticle dorylaimoid, smooth. Lip region variably offset, often marked by depression or constriction, with mostly separate, angular lips and protruding papillae. Amphid fovea cup- or funnel-like, with large aperture. Odontostyle strong, as long as lip region diameter or more, with aperture occupying up to one-half of its length. Guiding ring simple but distinct. Pharynx entirely muscular, gradually enlarging into the basal expansion that occupies ca one-half of the total neck length. Female genital system diovarian, with distinct pars refringens vaginae and transverse (occasionally longitudinal) vulva. Tail similar in the two sexes, conical, regularly curved ventrad (nearly straight in few cases). Spicules dorylaimid. Ventromedian supplements 5–20, spaced, lacking hiatus.
Relationships
As mentioned in the introductory section, Allodorylaimus is very similar to Eudorylaimus , from which it differs in the absence (vs presence) of pre-cloacal space (hiatus) in adult males; meanwhile, females are difficult to distinguish. In fact, when Andrássy (1986) created Allodorylaimus , he was aware that his proposal raised doubts about the identity of some parthenogenetic species of Eudorylaimus , stating (p. 9) ′It is possible that some of the species known recently in female forms only and ordered provisionally to the genus Eudorylaimus should later be transferred to Allodorylaimus ̍.
Allodorylaimus also resembles Aporcelinus . In fact, the separation of these two genera is not exempt from difficulty as it is based on subtle (but apparently significant) differences: nature of cuticle (the two layers with similar refractiveness vs without such similarity in Aporcelinus ), absence (vs presence) of a dorsal lobe at pharyngo-intestinal junction, uterine egg shell (smooth vs irregular, undulated), shape of sperm cells (spindle-shaped vs rounded to ovoid), and tail shape (very often regularly curved ventrad vs nearly always with a dorsal concavity and recurved dorsad).
Notes on its phylogeny
Allodorylaimus species only have in common the absence of a pre-cloacal space (hiatus) separating the pre-cloacal pair of genital papillae and the series of ventromedian supplements, which is herein regarded as a plesiomorphic condition. Thus, morphologically, the monophyly of this taxon is not supported by any relevant synapomorphy, much less by any autopomorphic trait. Its current geographical distribution suggests, however, a tentative origin of the group in the Laurasian supercontinent, in particular in Palearctic territories, with more recent and occasional dispersal events to other biogeographical regions. It belongs to a very diverse group of conical-tailed dorylaims classified under the family Qudsianematidae , whose monophyly is not supported by means of molecular data ( Holterman et al. 2008). Regarding molecular analyses, only one sequence each of 18S - and 28S -rRNA of A. andrassyi is currently available from GenBank – other deposited sequences were identified only at the genus level – and the molecular trees derived from their analysis show a closer relationship with members of the family Aporcelaimidae than with other qudsianematid taxa ( Holterman et al. 2008; Peña-Santiago et al. 2019).
Type species
A. uniformis ( Thorne, 1929) Andrássy, 1986
= Dorylaimus uniformis Thorne, 1929
= Eudorylaimus uniformis ( Thorne, 1929) Andrássy, 1959
= Dorylaimus acuticauda apud Steiner (1916) , nec de Man (1880)
Other valid species
A. aljabaranus Quijano, Peña-Santiago and Jiménez-Guirado, 1991
A. allgeni ( Andrássy, 1958) Andrássy, 1986
= Dorylaimus allgeni Andrássy, 1958
= Eudorylaimus allgeni ( Andrássy, 1958) Andrássy, 1959
= Dorylaimus carteri apud Allgén (1929) , nec Bastian (1865)
A. alpinus ( Steiner, 1914) Andrássy, 1986
= Dorylaimus alpinus Steiner, 1914
= Eudorylaimus alpinus ( Steiner, 1914) Andrássy, 1959
= Eudorylaimus sp. 3 apud Loof (1961)
A. andrassyi ( Meyl, 1955) Andrássy, 1986
= Dorylaimus andrassyi Meyl, 1955
= Eudorylaimus andrassyi ( Meyl, 1955) Andrássy, 1959
A. bokori (Andrássy, 1959) Andrássy, 1986
= Dorylaimus bokori Andrássy, 1959
= Eudorylaimus bokori (Andrássy, 1959) Andrássy, 1959
A. digiturus ( Thorne, 1939) Andrássy, 1986
= Dorylaimus digiturus Thorne, 1939
= Eudorylaimus digiturus ( Thorne, 1939) Andrássy, 1959
A. elytrigii Gagarin and Serikova, 2007
= Eudorylaimus andrassyi apud Tjepkema et al. (1971) , nec Meyl (1955) A. holdemani (Andrássy, 1959) Andrássy, 1986
= Dorylaimus holdemani Andrássy, 1959
= Eudorylaimus holdemani (Andrássy, 1959) Andrássy, 1959 A. husmanni ( Altherr, 1972) Andrássy, 1986
= Eudorylaimus husmanni Altherr, 1972
A. kazirangus Baniyamuddin and Ahmad, 2011
A. kosambaensis Khan, Ahmad and Jairajpuri, 1995
A. lindbergi ( Andrássy, 1960) Gagarin, 1997
= Eudorylaimus curvicaudatus Eliava, 1968
= Allodorylaimus rarus Gagarin, 1999a
A. paragranuliferus Quijano, Peña-Santiago and Jiménez-Guirado, 1991 A. parasimilis ( Kreis, 1963) Andrássy, 1986
= Dorylaimus parasimilis Kreis, 1963
= Eudorylaimus parasimilis ( Kreis, 1963) Andrássy, 1969 A. piracicabensis ( Lordello, 1955) Andrássy, 1986
= Dorylaimus piracicabensis Lordello, 1955
= Eudorylaimus piracicabensis ( Lordello, 1955) Andrássy, 1959 A. robustus ( Thorne, 1974) Andrássy, 1986
= Eudorylaimus robustus Thorne, 1974
A. santosi ( Meyl, 1957) Andrássy, 1986
= Dorylaimus santosi Meyl, 1957
= Eudorylaimus santosi ( Meyl, 1957) Andrássy, 1959
A. septentrionalis ( Kreis, 1963) Andrássy, 1986
= Dorylaimus septentrionalis Kreis, 1963
= Eudorylaimus septentrionalis ( Kreis, 1963) Andrássy, 1969 A. tarkoenensis (Andrássy, 1959) Andrássy, 1986
= Eudorylaimus tarkoenensis Andrássy, 1959
= Dorylaimus sp. apud Andrássy (1952)
A. thymophilus Quijano, Peña-Santiago and Jiménez-Guirado, 1991 A. tuccitanus sp. nov.
A. vallus Khan, Ahmad and Jairajpuri, 1995
Key to species identification
1a – Caudal region straight, dorsally first convex and then showing a weak but distinguishable concavity ........................................................................................................................ 2
1b – Caudal region more or less regularly curved ventrad ...................................................... 4
2a – Odontostyle 19–21 µm long ....................................... ....................................... kosambaensis
2b – Odontostyle 23 µm long or more ............................................................................................. 3
3a – Lip region 21 µm wide .............................................................................................. holdemanni
3b – Lip region 17–19 µm wide ................................................. ................................................. vallus
4a – Lip region continuous with the adjacent body ................................................................... 5
4b – Lip region offset from the adjacent body .............................................................................. 7
5a – Odontostyle ca 12 µm long; longer tail (54 µm, c = 24, c̾ = 1.9). ........ ........ digiturus
5b – Odontostyle up to 17 µm long; shorter tail (<48 µm, c> 27, c̾ up to 1.62) .......... 6
6a – Body 1.16–1.34 mm long, pharyngeal expansion occupying 43–49% of the total neck length, spicule 39 µm long .......................................................................... tarkoenensis
6b – Body 1.51–1.69 mm long, pharyngeal expansion occupying 37–39% of the total neck length, spicule 55–59 µm long ...................................................... tuccitanus sp. nov.
7a – Larger general size, body 3.28 mm long .............................................................. parasimilis
7b – Smaller general size, very exceptionally exceeding 3.0 mm long, reaching 3.07 mm ............................................................................................................................................... 8
8a – Longer female tail (more than 100 µm, c̾> 3) ...................................................... lindbergi
8b – Shorter female tail (up to 80 µm, c̾ <2.5) ............................................................................. 9
9a – Odontostyle much longer (1.3–1.5 times) than lip region diameter ........................ 10
9b – Odontostyle equal to or hardly longer (up to 1.3 times) than lip region diameter ............................................................................................................................................. 11
10a – Lip region offset by deep constriction and about 16 µm wide, tail strongly curved ventrad, five ventromedian supplements .................................. .................................. bokori
10b – Lip region offset by weak constriction and 20–22 µm wide, tail slightly curved ventrad, 13–14 ventromedian supplements ........................... ........................... kazirangus
11a – Only one ventromedian supplement lying within the range of the spicules and widely separated from the others ................................................................ paragranuliferus
11b – Ventromedian supplements with another pattern, more or less regularly spaced ................................................................................................................................................ 12
12a – Abundant cuticular irregularities (wrinkles) around the vulva ........ ........ thymophilus 12b – No cuticular irregularity around the vulva .......................................................................... 13
13a – Vulva longitudinal .......................................................................................................................... 14
13b – Vulva transverse ............................................................................................................................. 15
14a – More slender female body (a = 45–52); tail of both sexes strongly curved ventrad .. ......................................................................................................................................... septentrionalis
14b – Less slender female body (a <35); tail of both sexes weakly curved ventrad .............. ................................................................................................................................................. ferrisorum
15a – Larger general size, body length more than 1.8 mm long, exceptionally less ..... 16
15b – Smaller general size, body length only exceptionally more than 1.8 mm long ..................................................................................................................................................... 20
16a – Caudal region bearing abundant saccate bodies ....................... ....................... husmanni
16b – Caudal region lacking saccate bodies ................................................................................... 17
17a – Odontostyle 24–29 µm long ........................................................................................ uniformis
17b – Odontostyle up to 23 µm long ................................................................................................ 18
18a – More posterior vulva (V = 52); only one ventromedian supplement lying within the range of spicules ......................................................... ......................................................... alpinus
18b – More anterior vulva (V <50); two to four ventromedian supplements lying within the range of spicules .................................................................................................................... 19
19a – Odontostyle 17.5 µm long, length almost equal to the lip region diameter; caudal region weakly curved ventrad and bearing a short hyaline portion ..... ..... andrassyi
19b – Odontostyle 21–23 µm long, length 1.2–1.3 times the lip region diameter; caudal region strongly curved ventrad and bearing a long hyaline portion .............. elytrigii
20a – Female up to 1.05 mm long ...................................................................................................... 21
20b – Female more than 1.20 mm long ............................................................................................ 22
21a – Caudal region comparatively shorter (c̾ = 1.0–1.1); male bearing 18 ventromedian supplements .............................................................................................................. piracicabensis
21b – Caudal region comparatively longer (c̾ = 1.7–2.2); male bearing 13–15 ventromedian supplements ........................................................ ........................................................ santosi
22a – Shorter tail (20–30 µm, c = 47–59); male caudal region strongly curved ventrad, hook-like ............................................................. ............................................................. meridianus
22b – Longer tail (> 35 µm, c <45); male caudal region slightly curved ventrad, not hook-like ........................................................................................................................................................ 23
23a – Neck more than 400 µm long; spicules 85 µm long; only one ventromedian supplement within the range of spicules ....................................... ....................................... robustus
23b – Neck less than 400 µm long; spicules less than 75 µm long; two or three ventromedian supplements within the range of spicules ................................................................ 24
24a – Lip region offset by depression; longer caudal region (c̾ = 1.3–2.1) with finely rounded tip; spicules 52–55 µm long .................................................................. aljabaranus
24b – Lip region offset by deep constriction; shorter caudal region (c̾ = 1) and with acute tip; spicules 60 µm long ...................................................................................................... allgeni
Table 2 View Table 2 provides a compendium of the main morphometric features of known populations/specimens of each species, together with its geographical origin and the corresponding bibliographic reference.
Comments on some species
Allodorylaimus kosambiensis . Several traits of this species, especially the presence of a distinct dorsal concavity of caudal region in both sexes, resemble the pattern found in Aporcelinus representatives; therefore, the hypothesis that it belongs to this genus should not be discarded.
Allodorylaimus holdemanni . It is probably the most widely distributed species of the genus ( Table 1 View Table 1 ; see also other records compiled by Peña-Santiago 2021), but available information about this taxon is heterogeneous and not always coincident. Thus, Andrássy̍s (1959a) original description shows a pattern (lip region offset by deep constriction and with separate lips, strong odontostyle with aperture occupying almost one-half of the total length, and female tail with a slight dorsal concavity) very similar to that found in Aporcelinus species. Converserly, Zullini̍s (1970) illustrations present a nearly straight tail with regularly convex dorsal side.
Allodorylaimus lindbergi . Available information about this species is not consistent, and doubt persists regarding its true identity. Andrássy (1960) originally described it on the basis of four females from freshwater habitats in Afghanistan, and characterised it by, among other features, 2.17–2.42 mm long body, odontostyle 18–19 µm long, pars refringens vaginae absent of weakly developed, slightly anterior vulva ( V = 44–46), and tail conical elongate (c = 20–24, c ̍ = 3.2–3.7) and strongly curved ventrad, almost hook-like . Altherr (1972) recorded 10 females of this species from two locations, also in freshwater habitats, in Sweden . These females displayed smaller general size (body length 1.30–2.10 vs 2.17– 2.42 mm) and less slender body (a = 24–33 vs 33–41), but are similar to the type material in their remaining morphometrics. Thus, doubt exists about its true identity. Gagarin (1992) studied five females from Taimyr Peninsula , which were appreciably smaller than those of the type population (body 1.55–1.85 vs 2.17–2.42 mm long) and with comparatively shorter tail (c´ = 2.1–2.4 vs c̾ = 3.2–3.7), two remarkable differences . Eliava (1968) described Eudorylaimus curvicaudatus from a Siberian (Komi ASSR) freshwater habitat . Although the original description of this species is laconic, basic morphometrics and other relevant traits (for instance, pars refringens vaginae slightly sclerotised) are similar to those of type specimens. Andrássy (1986) regarded it as a junior synonym of E. lindbergi , an action that might be justified. Gagarin (1999a) described Allodorylaimus rarus from Siberian freshwater habitats on the basis of 11 females and only one male. The females of this species showed wide ranges in most of their morphometrics (body 1.61–2.37 mm long, V = 39–50, tail 70– 123 µm long, c = 15–23, c̾ = 2.2–3.7); meanwhile, the only male displayed a significantly shorter tail (60 µm long, c = 29, c̾ = 1.3) than females, and five irregularly spaced ventromedian supplements, two of them within the range of spicules . The same author (Gagarin 1999ab) regarded A. rarus as identical to E. lindbergi as the morphometrics of females of the two species were very similar, and transferred the latter to Allodorylaimus . Nevertheless, Gagarin̍ s (1999b) action is questionable. First, females of A. rarus present ′ pars refringens vaginae consisting of two well developed and triangular sclerotizations̍ (confirmed in Gagarin̍s original Figure 2 View Figure 2 (g)), whereas members of the type population of E. linbergi lack pars refringens vaginae or this is weakly developed (see above). Second, the significant difference observed in tail length of females and male of A. rarus suggests that they might not be conspecific. In conclusion, there are reasons to doubt (i) the synonymy of E. linbergi and A. rarus , (ii) the transfer of E. lindbergi to Allodorylaimus , and (iii) whether females and males of A. rarus belong to the same species.
Allodorylaimus piracicabensis . This species was described by Lordello (1955) on the basis of only two females, and the author did not provide much information about it. Later, Monteiro (1970) studied 10 females and one male that differ from those described by Lordello in their slightly larger size (body 1.17–1.34 vs 0.92–0.94 mm), less slender body (a = 22–28 vs a = 16–19), and longer odontostyle (18–20 vs 13–18 µm). Thus, doubt persists on whether these two groups of specimens belong to the same species. Remarkably, the only male known combines a comparatively short body with a high number (18) of ventromedian supplements.
Allodorylaimus rarus . See detailed explanation under A. lindbergi .
Allodorylaimus vallus . A few morphological traits of this species support its tentative assignment to the genus Aporcelinus – for instance, its strong odontostyle with aperture occupying ca one-half of its total length, and female tail with a weak (but appreciable) dorsal concavity. Furthermore, fig. 2(e) of Khan et al. (1995) suggests that a dorsal lobe might be present at the pharyngo-intestinal junction, as occurs in representatives of Aporcelinus .
| V |
Royal British Columbia Museum - Herbarium |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |