
Australocoelia palmata ( Morris & Sharpe, 1846 )
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4683.4.3 |
|
publication LSID |
lsid:zoobank.org:pub:8AB14BD4-AE9C-4FDA-9326-49E6BB4DA38A |
|
DOI |
https://doi.org/10.5281/zenodo.5929635 |
|
persistent identifier |
https://treatment.plazi.org/id/03A40F43-FFB3-4C23-FF39-F8FDFD05FC60 |
|
treatment provided by |
Plazi |
|
scientific name |
Australocoelia palmata ( Morris & Sharpe, 1846 ) |
| status |
|
Australocoelia palmata ( Morris & Sharpe, 1846)
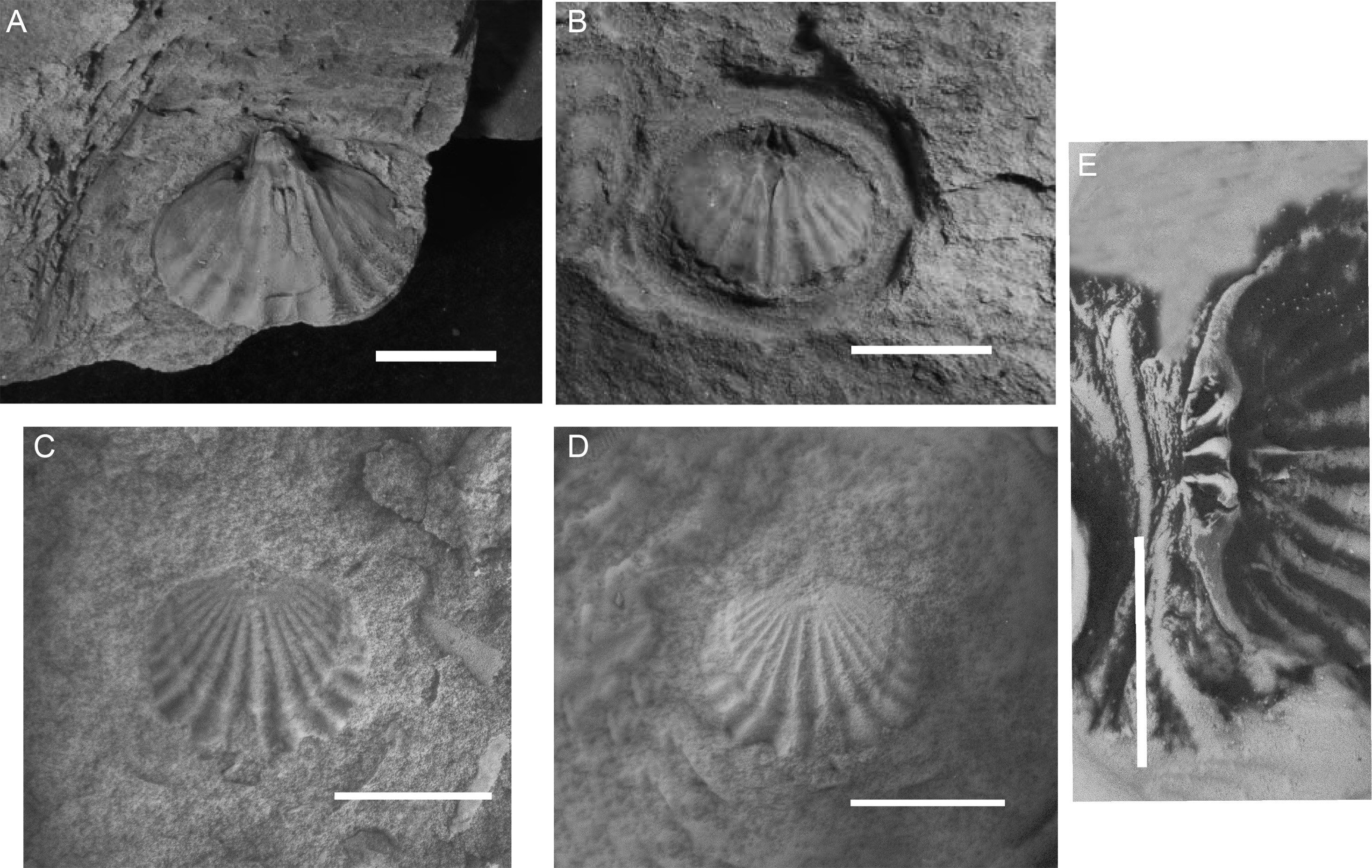
Figure 3 View FIGURE 3
1842 Terebratula peruviana d’Orbigny , v. 3, p. 36, plate 2, fig. 22-25.
1846 Atrypa palmata Morris & Sharpe , plate 26, fig. 3a-d.
1856 Orthis palmata (Morris & Sharpe) . Sharpe, plate 26, figs. 6-10.
1861 Orthis aymara Salter , plate 4, fig. 14.
1892 Leptocoelia flabellites [non Conrad] Ulrich, p. 60, plate 4, figs. 9-13.
1897 Vitulinia pustulosa [non Hall] Kayser, plate 9, figs. 6, 7, 9.
1897 Leptocoelia flabellites [non Conrad] Katzer, 2 (2): 231.
1903 Leptocoelia flabellites [non Conrad] Reed, plate 23, fig. 10.
1913 Leptocoelia flabellites [non Conrad] Clarke, plate 22, figs. 13-24.
1934 Leptocoelia flabellites [non Conrad] Méndez-Alzola, plate 7, figs. 4-10.
1956 Australocoelia tourteloti Boucot & Gill , plate 126, fig. 3, 4, 7-9.
1977 Australocoelia peruviana (d’Orbigny) Copper , 21 (3): 175.
1977 Australocoelia palmata (Morris & Sharpe) Isaacson , plate 1, Figs 10-25.
1993 Australocoelia palmata (Morris & Sharpe) Isaacson , plate 3, figs. 10-25.
Diagnosis: Australocoelia with posteriorly narrow costae, varying from 12 to 15, with 5 to 7 in each flank of each valve, in addition to those in the dorsal sulcus and the ventral fold; Costa gradually widening as it approaches the anterior commissure, becoming subrounded, angled and slightly curved. Small interspace when compared to costa size.
Material: Interior ventral mold: UNIRIO 0061-BQ; UNIRIO 0080-BQ; UNIRIO 0099-BQ; UNIRIO 0101- BQ; UNIRIO 0115-BQ; UNIRIO 0129-BQ; UNIRIO 0152-BQ; UNIRIO 0183-BQ; UNIRIO 0184-BQ; UNIRIO 0185-BQ; UNIRIO 0196-BQ; UNIRIO 0212-BQ; UNIRIO 0230 -BQ; UNIRIO 0509-BQ; UNIRIO 0530-BQ; UNIRIO 0533-BQ; UNIRIO 0503-BQ; UNIRIO 0550-BQ; UNIRIO 0566-BQ; UNIRIO 0569-BQ; UNIRIO 0574- BQ; UNIRIO 0577-BQ; UNIRIO 0579-BQ; UNIRIO 0581-BQ; UNIRIO 0595-BQ; UNIRIO 0614-BQ; UNIRIO 0616-BQ; UNIRIO 0639-BQ; UNIRIO 0689-BQ; UNIRIO 0703-BQ; Interior dorsal mold: UNIRIO 0085-BQ; UNIRIO 0086-BQ; UNIRIO 0092-BQ; UNIRIO 0099-BQ; UNIRIO 0129-BQ; UNIRIO 0166-BQ; UNIRIO 0170- BQ; UNIRIO 0183-BQ; UNIRIO 0192-BQ A e B; UNIRIO 0196-BQ; UNIRIO 0211-BQ; UNIRIO 0215-BQ; UNIRIO 0526-BQ; UNIRIO 0515-BQ; UNIRIO 0516-BQ; UNIRIO 0537-BQ; UNIRIO 0557-BQ; UNIRIO 0564- BQ; UNIRIO 0546-BQ; UNIRIO 0547-BQ; UNIRIO 0566-BQ; UNIRIO 0568-BQ; UNIRIO 0572-BQ; UNIRIO 0573-BQ; UNIRIO 0575-BQ; UNIRIO 0578-BQ; UNIRIO 0579-BQ; UNIRIO 0581-BQ; UNIRIO 0582-BQ; UNIRIO 0583-BQ; UNIRIO 0585-BQ; UNIRIO 0590-BQ; UNIRIO 0619-BQ; UNIRIO 0620-BQ; UNIRIO 0636- BQ; UNIRIO 0637-BQ; UNIRIO 0640-BQ A e B; UNIRIO 0641-BQ; UNIRIO 0690-BQ; UNIRIO 0702-BQ; UNIRIO 0730-BQ; UNIRIO 0738-BQ; UNIRIO 0754-BQ; Exterior ventral mold: UNIRIO 0082-BQ; UNIRIO 0092-BQ; UNIRIO 0142-BQ; UNIRIO 0156-BQ; UNIRIO 0165-BQ; UNIRIO 0177-BQ; UNIRIO 0206-BQ; UNIRIO 0210-BQ; UNIRIO 0212-BQ; UNIRIO 0510-BQ; UNIRIO 0527-BQ; UNIRIO 0557-BQ; UNIRIO 0567- BQ; UNIRIO 0537-BQ; UNIRIO 0579-BQ; UNIRIO 0589-BQ; UNIRIO 0599-BQ; UNIRIO 0617-BQ; Exterior dorsal mold: UNIRIO 0156-BQ; UNIRIO 0165-BQ; UNIRIO 0167-BQ; UNIRIO 0170-BQ; UNIRIO 0180-BQ; UNIRIO 0196-BQ; UNIRIO 0220-BQ; UNIRIO 0509-BQ; UNIRIO 0510-BQ; UNIRIO 0554-BQ; UNIRIO 0565- BQ; UNIRIO 0565-BQ; UNIRIO 0540-BQ; UNIRIO 0569-BQ; UNIRIO 0581-BQ; UNIRIO 0596-BQ; UNIRIO 0607-BQ; UNIRIO 0617-BQ; UNIRIO 0680-BQ; UNIRIO 0685-BQ; UNIRIO 0737-BQ; UNIRIO 0746-BQ; Composite ventral mold: UNIRIO 0210-BQ; Composite dorsal mold: UNIRIO 0062-BQ; Ventral interior countermold: UNIRIO 0619-BQ; Dorsal interior counter-mold: UNIRIO 0092-BQ; UNIRIO 0142-BQ; UNIRIO 0154- BQ; Ventral exterior counter-mold: UNIRIO 0205-BQ; Dorsal exterior counter-mold: UNIRIO 0234-BQ; Hinged valves exterior counter-mold: UNIRIO 0163-BQ; UNIRIO 0165-BQ; UNIRIO 0198-BQ; UNIRIO 0206- BQ; Hinged valves interior mold: UNIRIO 0142-BQ; UNIRIO 0156-BQ; UNIRIO 0164-BQ; UNIRIO 0167-BQ; UNIRIO 0169-BQ; UNIRIO 0170-BQ; UNIRIO 0172-BQ; UNIRIO 0177-BQ; UNIRIO 0180-BQ; UNIRIO 0206- BQ; UNIRIO 0207-BQ; UNIRIO 0213-BQ; UNIRIO 0573-BQ; Hinged valves exterior mold: UNIRIO 0163-BQ; UNIRIO 0168-BQ;
Occurrence: Rivadavia Farm 01: Railway, inside Rivadavia Farm, 5.3 km from the farm entrance. Railroad section of Ponta Grossa—Paranaguá, km 222, Lineu Amaral Station, Municipality of Ponta Grossa, PR. This outcrop was located in an area of the former Rivadavia Farm, which was later dismembered, situated in an area currently called Santa Helena Farm.
Rail curve 02: Central section of the Paraná railroad, section Uvaranas—Apucarana, subsection Uvaranas—Pe-riquitos, approximately 2 km from the outcrop Rail curve 01, in the next curve of the railroad, Municipality of Ponta Grossa , PR .
Rail curve 01: Central section of the Paraná railroad, section Uvaranas—Apucarana, subsection Uvaranas—Pe-riquitos, km 25, Vila Baraúna, about 800 m right of the intersection with Monteiro Lobato Avenue, exit to Castro, Municipality of Ponta Grossa , PR .
Jaguariaíva 03: Classic railroad cut, railway branch of Jaguariaíva—Arapoti between km 2.2 and 6.6, Municipality of Jaguariaíva ( SIGEP 65). Located 360 m from Jaguariaíva 02, on the railway towards Jaguariaíva, about 2.8 km from José Gonçalves de Jaguariaíva Square (PR), after crossing the bridge over the river. Turn left at the first cloverleaf in the freeway toward Lagoão.
Boa Vista Farm 01: Outcrop in the gravel pit, at Boa Vista Farm, 7.1 km from the farm entrance. Farm located 4.3 km from the entrance bridge of Ponta Grossa city on the BR.376, and 8.1 km from the Cathedral Square of the city of Ponta Grossa, Ponta Grossa, PR.
Jaguariaíva 01: Classic outcrop of the railroad cut, railway branch of Jaguariaíva—Arapoti between km 2.2 and 6.6, Municipality of Jaguariaíva ( SIGEP 65). Collecting point to the right of the intersection with RFFSA, 2.5 km from José Gonçalves Square in the city of Jaguariaíva (PR), after crossing the bridge over the river. Turn left at the first cloverleaf in the freeway toward Lagoão.
Description—Exterior: Shell of subcircular outline, moderate size, unequally biconvex. Suberect beak. Posteriorly narrow costae, widening gradually as it approaches the anterior commissure, acquiring a sub-rounded, angular and slightly curved aspect. Reduced interspace when compared to costa size. Concentric growth lines, and crenulated anterior margin. Ventral groove marked by a single costa, with a pair marking the fold of the opposite valve. The number of costae varies from 12 to 15, with 5 to 7 on each flank of each valve, plus the ones corresponding to the ventral groove and the dorsal fold.
Ventral interior: Stout triangular hinge teeth, fine in the base widening towards the apex. Pair of broad diductor muscle scars, anteriorly rounded narrowing as it approaches the posterior beak. Flabellate adductor scar. Median septum extending anteriorly, dividing both muscle scars.
Dorsal interior: Cardinal process in a mound-like shape (unilobed), terminally swollen. Crural plates lateral to the cardinal process. Dental sockets triangular in shape, broad anteriorly, narrowing as it approaches the apex. Median septum extending anteriorly about one-third the length of the valve.
Discussion: Australocoelia palmata ’s occurrence in the Devonian of Brazil was considered restricted to the Paraná Basin until the first record made by Gama Jr (2008) in the western margin of the Parnaíba Basin.
The first recognition of Devonian age of the rocks from the Pimenteira Formation was made by Dr. Kenneth E. Caster, based on a field trip in 1947 with Dr. Llewellyn I. Price. The collected material served as object of study for two dissertations in the University of Cincinnati entitled “ Devonian fauna in the concretionary Picos Member, Pimenteira Formation (Lower Devonian) Piauí, Brazil ” by David Ford in 1965; and “ Some Devonian fossils from the State of Piauí, Brazil ” by Mario Suárez Riglos in 1967 ( Lima & Ponciano, 2017). From all the studied samples, Suárez-Riglos (1967) mentioned the possible occurrence of Australocoelia tourteloti , due to the external resemblance of the molds with the so-called characteristic “Austral” species of Boucot & Gill (1956). However, the lack of interior molds did not allow a precise confirmation of its presence.
During the fieldwork coordinated by LECP’s (Laboratório de Estudos de Comunidades Paleozoicas) members, new specimens were collected along the eastern margin of this same basin, as pointed out by Ponciano et al. (2012a), in which it was again indicated with doubts, the possible occurrence of this species in two different localities: Picos 2 outcrop (BR-316/Km 318) and Mucambo outcrop (PI-466/Km 12), in the State of Piauí. The analysis of Australocoelia palmata ’s specimens from the Parnaíba Basin, collected in Riachão outcrop (Picos-Itainópolis road BR-020, right after Riachão village, 27 Km from Picos, in Itainópolis city) compared with the ones from the Paraná Basin, made it clear that they would belong in the same species due to the similarity between the diagnostic characters, despite the different kinds of preservation.
The material from the eastern margin of the Parnaíba Basin consists of exterior and interior dorsal molds, both presenting subcircular outlines and subrounded costae that gradually increase in size from the beak towards the anterior commissure. There is an average of five costae in each flank, in addition to the pair related to the fold, and a mound-like/unilobed cardinal process identical to those identified by Gama Jr (2008), that matches A. palmata .
Its presence in Pimenteira Formation’s Middle Devonian (Picos Member) could be explained by paleogeographic factors, such as the marine transgression that occurred during Eifelian/Givetian boundary establishing a connection between the Paraná and Parnaíba basins, as discussed by Copper (1977), Melo (1988), Gama Jr (2008) and Grahn et al. (2016), creating a passageway in which this species could have made the migration.
Its presence was also confirmed in the Alto-Garças sub-Basin (Paraná Basin) by Kunzler (2012, fig. 19 D-F), in the following localities: Rondonópolis; São Jerônimo’s Hill; Véu da Noiva’s Waterfall; River Coxipó; Chapada dos Guimarães and an unknown locality from Chapada dos Guimarães, State of Mato Grosso; confirming the taxon’s wide distribution inside the Paraná Basin.
Besides the Paraná and Parnaíba basins, another occurrence is known for the Brazilian Devonian strata. Boucot et al. (2001) studied samples from Morro Vermelho and Morro do Indio localities, north of Paranatinga town, Parecis Basin, State of Mato Grosso. The fossils and the lithology resemble the Lower Devonian of Chapada dos Guimarães, suggesting that these two regions were formerly continuous. In both areas were found Malvinokaffric elements, and among them, the endemic Australocoelia palmata . According to the authors, the studied material from the Parecis Basin has the typical characters of the species.
Another Devonian occurrence of Australocoelia was reported by Barretto (1968) from the Upper Devonian Inajá Formation (Jatobá Basin, State of Pernambuco), but without illustrations or descriptions. Later, Muniz (1976) and Copper (1977) reexamined the material, but due to its poorly preserved state, it could only be ascribed as a Rhynchonellida .
A comparison between the species A. polyspera ( Gill, 1950) and A. palmata highlights the fact that the former has a more orbicular outline and ornamentation, composed of more rounded and numerous costae and reduced interspace (fide in Gill, 1950, plate figs. 26–28, 38).
In spite of morphological divergences, the wide distribution of the genus in the austral regions during the Devonian, not only in Malvinokaffric cold water, but also in Australian (non Malvinokaffric) warm water with its climate similar to the boreal region strongly supports Boucot & Gill’s (1956) affirmation which discusses Australocoelia ’s main distribution motive as being an eurythermal form that allows larger tolerance to temperature variations.
This ecological aspect can also be seen through the comparison between specimens collected in the Paraná and Parnaíba basins, in the differences related to temperature, being an environment of cold water in the former and of warm water in the latter ( Copper, 1977), besides the transgressive peak, that caused the communication between both basins during the Eifelian/Givetian boundary. A. palmata ’s presence on both western and eastern side of the Pimenteira Formation (Picos Member) extends the range of occurrence and reinstates the idea of a eurythermal form of this genus. Such environmental differences found in the Ponta Grossa and Pimenteira formations is justified by Fonseca & Melo (1987) as a rearrangement of the ocean circulation patterns during Middle Devonian, reaching its climax with the transgressive event recorded in the Parnaíba and Amazonas basins (Pimenteira and Maecuru formations respectively) with warmer water flows. This later reached the Paraná Basin, causing its massive extinction event during early Givetian.
Another aspect that supports the eurythermal hypothesis of the genus is the presence of Australocoelia palmata in the Devonian of the Zorritas Formation (Northern Chile), as pointed out by Isaacson et al. (1985). Boucot et al. (1995) discussed the Zorritas Formation as representing a boundary of a mixing area between the Malvinokaffric and extra-Malvinokaffric Realms, with elements from cool and warm conditions, like the typical Malvinokaffric Australocoelia , and the immigrant Tropidoletpus from the Old World Realm. Rearrangements of ocean circulation during Middle Devonian, associated with sea level rise and expansion, created marine passageways that helped to establish connections between South America, South Africa and North America epicontinental seas ( Isaacson & Perry, 1977b; Fonseca & Melo, 1987; Barrett & Isaacson, 1988).
Geographic, Lithostratigraphic, Chronostratigraphic Distribution: Falkland Islands (Fox Bay Formation); Brazil (Ponta Grossa, São Domingos and Pimenteira formations and Chapada Group); South Africa (Bokkeveld Group); Argentina (Talacasto, Punta Negra, Cerro Piedras, Copo, Caburé and Rincón Formations); Bolivia (Icla, Belém, Huamampampa, Sica Sica and Gamoneda Formations); Peru (Lamba Formation); Uruguay (Cordobés Formation); Venezuela (Caño Grande Formation).
| BR |
Embrapa Agrobiology Diazothrophic Microbial Culture Collection |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |