
Australocoelia boucoti, Rezende & Machado & Ponciano, 2019
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4683.4.3 |
|
publication LSID |
lsid:zoobank.org:pub:8AB14BD4-AE9C-4FDA-9326-49E6BB4DA38A |
|
persistent identifier |
https://treatment.plazi.org/id/EE5CD9FB-7112-4976-ADE7-F114914EBAC8 |
|
taxon LSID |
lsid:zoobank.org:act:EE5CD9FB-7112-4976-ADE7-F114914EBAC8 |
|
treatment provided by |
Plazi |
|
scientific name |
Australocoelia boucoti |
| status |
sp. nov. |
Australocoelia boucoti , n. sp.
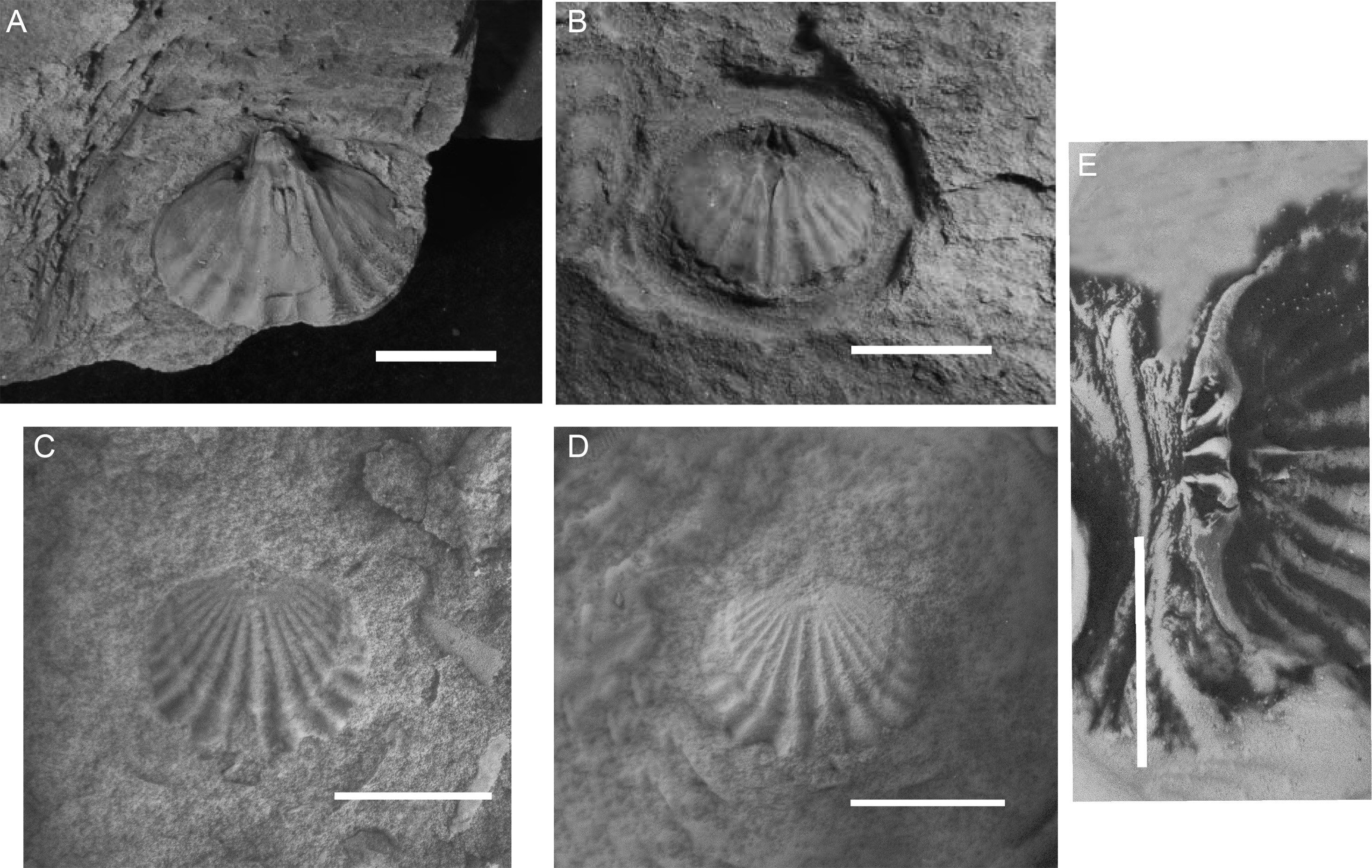
Figure 4 View FIGURE 4
Holotype: UNIRIO 0531-BQ ( Fig. 4.A View FIGURE 4 )
Paratype: UNIRIO 0506-BQ
Etymology: Species dedicated to Arthur James Boucot, one of the paleontologists responsible for the description of Australocoelia .
Type horizon: Railroad section of Ponta Grossa—Paranaguá, Km 222, Lineu Amaral Station, Municipality of Ponta Grossa.
Type locality: Ponta Grossa Formation
Material: Interior ventral mold: UNIRIO 0502-BQ; UNIRIO 0531-BQ; UNIRIO 0749-BQ; Exterior ventral mold: UNIRIO 0506-BQ; UNIRIO 0571-BQ UNIRIO 0738-BQ; UNIRIO 0753-BQ; UNIRIO 0747-BQ; Interior dorsal mold: UNIRIO 641-BQ A e B; UNIRIO 0750-BQ; UNIRIO 0752-BQ A e B; UNIRIO 0751-BQ; Exterior dorsal mold: UNIRIO 0571-BQ; UNIRIO 0748-BQ
Diagnosis: Australocoelia with sharply incised costae, which develops in a continuous way from the umbo, not getting rounded as it approaches the anterior margin, without a subrounded aspect and bigger interspace compared to costa size.
Description. Exterior: Subcircular shell of moderate size and unequally biconvex. Costae sharply incised, which extends in a continuous way from the umbo towards the anterior margin without getting subrounded as it approaches the commissure. Bigger interspace compared to costa size. Anterior crenulate commissure and suberect beak. Ventral groove marked by a single costa, with a pair marking the dorsal fold in the opposite valve. The number of costae varies from 12 to 15, with 5 to 7 in each flank, plus the ones corresponding to the ventral groove and the dorsal fold.
Ventral interior: Subcircular shells, moderate size. Marked convexity, stout triangular teeth, fine in the base widening towards the apex. Elongated adductor muscle scar divided by a short median septum that extends until the anterior end of the muscular field. Well-marked costae, starting thin from the umbo, not getting angled as it approaches the anterior margin, with an incised form and not acquiring a sub-angled shape as it approaches the commissure. The number varies from 5 to 6 in each flank, plus the one that corresponds to the groove. Crenulate commissure.
Dorsal interior: Cardinal process in a mound-like shape (unilobed) terminally swollen. Thin crural plates, lateral to the cardinal process. Dental sockets form a sulcus on the posterior margin of the valve, in a triangular shape, broad anteriorly narrowing closer to the apex. Median septum extending from the cardinalia towards the commissure, reaching about half the length of the valve.
Discussion: Copper (1977) indicated the presence of a new species, less common, also belonging to the genus Australocoelia that occurred in the Tibagi Member (Eifelian/Givetian). It was recognized by its “sharply incised costae”. While discussing the matter Melo (1985), due to not only the absence of photos but also of samples, decided to keep the classification proposed by Boucot & Gill (1956).
Based on the morphological characters of the samples described above, it is possible to determine that the specimens found in the Ponta Grossa Formation were from a different species, and not Australocoelia palmata ( Morris & Sharpe, 1846) .
When analyzed and compared with the diagnosis of the type-species, it became clear that the new specimens from the Ponta Grossa Formation belonged to the genus Australocoelia , but in a different species. The main character that allowed the identification and separation of both species was the ornamentation, in which the pattern of costae and interspace found in the molds were also used by Boucot & Gill (1956), being considered diagnostic in the differentiation of Australocoelia , the Malvinokaffric A. palmata and the non Malvinokaffric A. polyspera from Australia.
As discussed by Reed (1925) regarding the specimens of Leptocoelia flabellites from the Southern Hemisphere (currently A. palmata ), the amount and variety of samples have different levels of preservation, making proper identification difficult.
Based on a taphonomic analysis, it is observed that there is a great variety among the Australocoelia samples, especially related to the layers preserved in each mold. Such variation is most evident through the costa thickness, which becomes gradually smaller as it gets more internalized. The thickness of the costae could influence taxonomic identification, but, in spite of the mold being found with a more inner layer, the most characteristic aspect of A. palmata , the angled and rounded aspect of the costa in its anterior margin remains preserved, allowing an abundant Malvinokaffric species to be rapidly recognized and identified, keeping clear its differences from A. boucoti n. sp.
Another taphonomic aspect that has shown itself troublesome was the presence of a wing-like structure in the right flank of A. boucoti ’s valves, a possible consequence of the rocks plasticity in which the organisms were fossilized, providing a peculiar asymmetry to some of the specimens. This taphonomic matter becomes a problem during the process of taxonomic identification of a new species, as mentioned by Machado (1999) about the problematic internal plate seen in bivalves from the genus Nuculites which, as consequence of tectonism or depositional deformations, destroyed the main characteristics of that structure considered by the author as one of the most important to differentiate the species within this genus.
The preservation quality of all studied specimens makes it difficult to classify each taxon. The comparison between A. palmata and A. boucoti helped to identify both species, based on the differences that exist on their diagnostic characters ornamentation and interspace ( Figure 5 View FIGURE 5 ). More specifically about ornamentation, it is worth mentioning that the costae are sharp and develop in a continuous way from the beak to the anterior margin in A. boucoti , not getting the common rounded aspect that defines A. palmata , regardless of possible taphonomic variations. The difference observed in the ornamentation pattern has as consequence a variation in the interspace pattern. A. boucoti has bigger interspaces when compared to A. palmata , which due to rounded costae has its interspaces diminished.
In spite of the difficulties related to the specimen’s preservation, it was possible to identify with certainty a new morphotype, whose main difference consists in the well-marked ornamentation which fits within the genus Australocoelia based on the same characters described by Savage et al. in Williams et al. (2000).
Accompanying the taphonomic, sampling and time averaging issues, some other ecological aspects can be raised, given the morphological differences identified in A. palmata and A. boucoti n. sp. Fluctuations in temperature, salinity and food supply can develop multiple phenotypes (ecophenotypes), in a different way than a subspecies, whose population becomes adapted to an ecologic niche. However, it is unusual that subspecies are found coexisting in the same geographic and ecologic region, also unlikely to occur in a single fossil fauna. Since both Australocoelia ’s species occur together ( Figure 5 View FIGURE 5 ), the idea of subspeciation is refuted ( Newell, 1948; Pfenning et al, 2010; Dielt, 2013; Nagel-Myers et al. 2018). However, since A. palmata and A. boucoti n. sp. were found in the same habitat and closer ecological niche, the possibility of competition cannot be disregarded. According to Alexander (1976), contemporaneous Paleozoic brachiopods would subdivide their feeding niches, in order to avoid competition.
Furthermore, the wide, cosmopolitan distribution of Australocoelia palmata in the Malvinokaffric Realm and non Malvinokaffric area, as discussed above, regarding the Parnaíba Basin ( Figure 3 View FIGURE 3 ; C-D) (Gama Jr, 2008; Ponciano et al. 2012a), with no variation in either valves, presenting in all scenarios the same morphological pattern in ornamentation, even under taphonomic variations, does not agree with a phenotypic plasticity. Especially when taken into account the discussion about homogeneous environment, also seen in the Malvinokaffric Realm, provided by Alexander (1976), and the fact that ecophenotype cannot typify entire populations, as advocated by Newell (1948); Pfenning et al. (2010) and Nagel-Myers et al. (2018). We here erect a new species of, so far, narrower distribution, and probably even more typical cold-water Malvinokaffric Australocoelia , entitled Australocoelia boucoti n. sp. Some assumptions can be made regarding its absence in other regions. As observed in Australocoelia polyspera which is restricted to the warmer Australian region, Australocoelia boucoti seems to be exclusive to cold water environments. Such a geographic range could be an explanation of its lower abundance, since Australocoelia palmata had broader tolerance to climatic shifts, being able to extract more resources and occupy different regions, as confirmed by its abundance, enormous distribution, and no morphological changes.
Occurrence: Rivadávia Farm 01: Railway, inside Rivadavia Farm, 5.3 km from the farm entrance. Railroad section of Ponta Grossa—Paranaguá, km 222, Lineu Amaral Station, Municipality of Ponta Grossa, PR. This outcrop was located in an area of the former Rivadavia Farm, which was later dismembered, situated in an area currently called Santa Helena Farm.
Jaguariaíva 1: Railroad section, railway branch of Jaguariaíva-Arapoti between km 2.2 and 6.6, Municipality of Jaguariaíva (SIGEP 65)
Stratigraphic distribution: Ponta Grossa Formation (Pragian—Emsian)


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |